

Cellular immediate-early gene expression occurs kinetically upstream of Epstein-Barr virus bzlf1 and brlf1 following cross-linking of the B cell antigen receptor in the Akata Burkitt lymphoma cell line
- PMID: 20861250
- PMCID: PMC2976389
- DOI: 10.1128/JVI.01415-10
Cellular immediate-early gene expression occurs kinetically upstream of Epstein-Barr virus bzlf1 and brlf1 following cross-linking of the B cell antigen receptor in the Akata Burkitt lymphoma cell line
Abstract
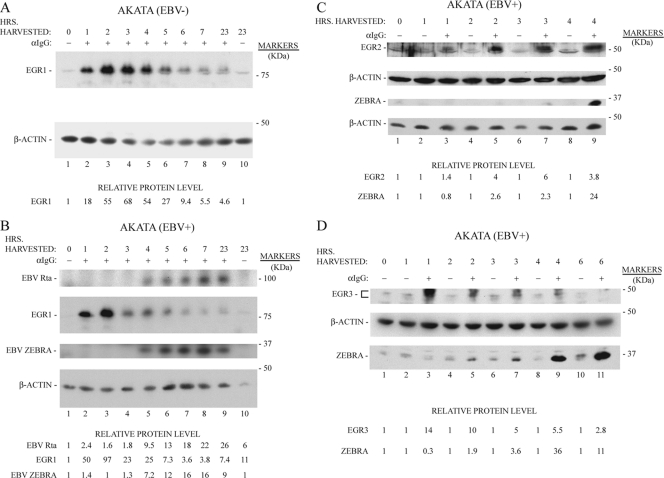
The Epstein-Barr virus (EBV) lytic activator genes bzlf1 and brlf1 are conventionally referred to as immediate-early (IE) genes. However, previous studies showed that the earliest expression of these genes was blocked by cycloheximide when the EBV lytic cycle was induced by histone deacetylase (HDAC) inhibitors and protein kinase C agonists. Anti-IgG activates a complex signal transduction pathway that leads to EBV lytic activation in the Akata cell line. Here we demonstrate that in Akata cells, where lytic cycle activation occurs very rapidly after anti-IgG treatment, de novo protein synthesis is also required for induction of bzlf1 and brlf1 expression. New protein synthesis is required up to 1.25 h after application of anti-IgG; bzlf1 and brlf1 mRNAs can be detected 1.5 h after anti-IgG. Five cellular IE genes were shown to be expressed by 1 h after addition of anti-IgG, and their expression preceded that of bzlf1 and brlf1. These include early growth response genes (egr1, egr2, and egr3) and nuclear orphan receptors (nr4a1 and nr4a3). These genes were activated by anti-IgG treatment of Akata cells with and without the EBV genome; therefore, their expression was not dependent on expression of any EBV gene product. EGR1, EGR2, and EGR3 proteins were kinetically upstream of ZEBRA and Rta proteins. Expression of EGR1, ZEBRA, and Rta proteins were inhibited by bisindolylmaleimide X, a selective inhibitor of PKC. The findings suggest a revised model in which the signal transduction cascade activated by cross-linking of the B cell receptor induces expression of cellular IE genes, such as early growth response and nuclear orphan receptor genes, whose products, in turn, regulate bzlf1 and brlf1 expression.
Figures









References
-
- Adamson, A. L., D. Darr, E. Holley-Guthrie, R. A. Johnson, A. Mauser, J. Swenson, and S. Kenney. 2000. Epstein-Barr virus immediate-early proteins BZLF1 and BRLF1 activate the ATF2 transcription factor by increasing the levels of phosphorylated p38 and c-Jun N-terminal kinases. J. Virol. 74:1224-1233. - PMC - PubMed
-
- Ben-Sasson, S. A., and G. Klein. 1981. Activation of the Epstein-Barr virus genome by 5-aza-cytidine in latently infected human lymphoid lines. Int. J. Cancer 28:131-135. - PubMed
-
- Bhaduri-McIntosh, S., and G. Miller. 2006. Cells lytically infected with Epstein-Barr virus are detected and separable by immunoglobulins from EBV-seropositive individuals. J. Virol. Methods 137:103-114. - PubMed
-
- Bhende, P. M., W. T. Seaman, H. J. Delecluse, and S. C. Kenney. 2004. The EBV lytic switch protein, Z, preferentially binds to and activates the methylated viral genome. Nat. Genet. 36:1099-1104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
