

A hierarchical phosphorylation cascade that regulates the timing of PERIOD nuclear entry reveals novel roles for proline-directed kinases and GSK-3beta/SGG in circadian clocks
- PMID: 20861372
- PMCID: PMC2957474
- DOI: 10.1523/JNEUROSCI.1586-10.2010
A hierarchical phosphorylation cascade that regulates the timing of PERIOD nuclear entry reveals novel roles for proline-directed kinases and GSK-3beta/SGG in circadian clocks
Abstract
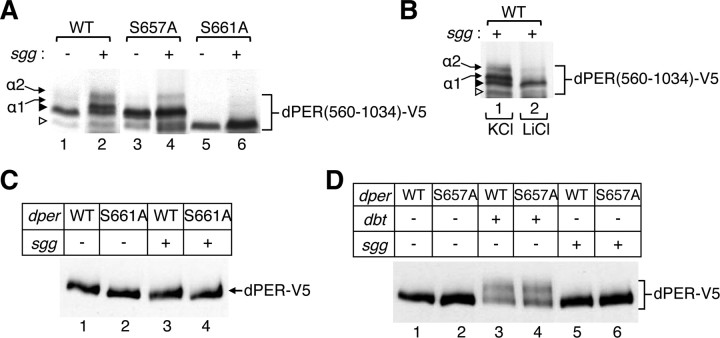
The daily timing of when PERIOD (PER) proteins translocate from the cytoplasm to the nucleus is a critical step in clock mechanisms underpinning circadian rhythms in animals. Numerous lines of evidence indicate that phosphorylation plays a prominent role in regulating various aspects of PER function and metabolism, including changes in its daily stability and subcellular distribution. In this report, we show that phosphorylation of serine 661 (Ser661) by a proline-directed kinase(s) is a key phospho-signal on the Drosophila PER protein (dPER) that regulates the timing of its nuclear accumulation. Mutations that block phosphorylation at Ser661 do not affect dPER stability but delay its nuclear entry in key pacemaker neurons, yielding longer behavioral rhythms. Intriguingly, abolishing phosphorylation at Ser661 also attenuates the extent of dPER hyperphosphorylation in vivo, suggesting the phosphorylated state of Ser661 regulates phosphorylation at other sites on dPER. Indeed, we identify Ser657 as a site that is phosphorylated by the glycogen synthase kinase GSK-3β (SHAGGY; SGG) in a manner dependent on priming at Ser661. Although not as dramatic as mutating Ser661, mutations that abolish phosphorylation at Ser657 also lead to longer behavioral periods, suggesting that a multi-kinase hierarchical phosphorylation module regulates the timing of dPER nuclear entry. Together with evidence in mammalian systems, our findings implicate proline-directed kinases in clock mechanisms and suggest that PER proteins are key downstream targets of lithium therapy, a potent inhibitor of GSK-3β used to treat manic depression, a disorder associated with clock malfunction in humans.
Figures





Similar articles
-
Two distinct modes of PERIOD recruitment onto dCLOCK reveal a novel role for TIMELESS in circadian transcription.J Neurosci. 2010 Oct 27;30(43):14458-69. doi: 10.1523/JNEUROSCI.2366-10.2010. J Neurosci. 2010. PMID: 20980603 Free PMC article.
-
A role for O-GlcNAcylation in setting circadian clock speed.Genes Dev. 2012 Mar 1;26(5):490-502. doi: 10.1101/gad.182378.111. Epub 2012 Feb 10. Genes Dev. 2012. PMID: 22327476 Free PMC article.
-
A key temporal delay in the circadian cycle of Drosophila is mediated by a nuclear localization signal in the timeless protein.Genetics. 2011 Jul;188(3):591-600. doi: 10.1534/genetics.111.127225. Epub 2011 Apr 21. Genetics. 2011. PMID: 21515571 Free PMC article.
-
Regulating a circadian clock's period, phase and amplitude by phosphorylation: insights from Drosophila.J Biochem. 2006 Nov;140(5):609-17. doi: 10.1093/jb/mvj198. Epub 2006 Sep 29. J Biochem. 2006. PMID: 17012288 Review.
-
Reversible protein phosphorylation regulates circadian rhythms.Cold Spring Harb Symp Quant Biol. 2007;72:413-20. doi: 10.1101/sqb.2007.72.048. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419299 Review.
Cited by
-
GSK-3 and CK2 Kinases Converge on Timeless to Regulate the Master Clock.Cell Rep. 2016 Jul 12;16(2):357-367. doi: 10.1016/j.celrep.2016.06.005. Epub 2016 Jun 23. Cell Rep. 2016. PMID: 27346344 Free PMC article.
-
High-Amplitude Circadian Rhythms in Drosophila Driven by Calcineurin-Mediated Post-translational Control of sarah.Genetics. 2018 Jul;209(3):815-828. doi: 10.1534/genetics.118.300808. Epub 2018 May 3. Genetics. 2018. PMID: 29724861 Free PMC article.
-
Glycogen synthase kinase is a regulator of the circadian clock of Neurospora crassa.J Biol Chem. 2012 Oct 26;287(44):36936-43. doi: 10.1074/jbc.M112.396622. Epub 2012 Sep 6. J Biol Chem. 2012. PMID: 22955278 Free PMC article.
-
Time-course RNASeq of Camponotus floridanus forager and nurse ant brains indicate links between plasticity in the biological clock and behavioral division of labor.BMC Genomics. 2022 Jan 15;23(1):57. doi: 10.1186/s12864-021-08282-x. BMC Genomics. 2022. PMID: 35033027 Free PMC article.
-
Glycogen synthase kinase-3 (GSK3): regulation, actions, and diseases.Pharmacol Ther. 2015 Apr;148:114-31. doi: 10.1016/j.pharmthera.2014.11.016. Epub 2014 Nov 27. Pharmacol Ther. 2015. PMID: 25435019 Free PMC article. Review.
References
-
- Akten B, Jauch E, Genova GK, Kim EY, Edery I, Raabe T, Jackson FR. A role for CK2 in the Drosophila circadian oscillator. Nat Neurosci. 2003;6:251–257. - PubMed
-
- Bae K, Edery I. Regulating a circadian clock's period, phase and amplitude by phosphorylation: insights from Drosophila. J Biochem. 2006;140:609–617. - PubMed
-
- Chang DC, Reppert SM. A novel C-terminal domain of Drosophila PERIOD inhibits dCLOCK:CYCLE-mediated transcription. Curr Biol. 2003;13:758–762. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases