

Two coordinately regulated homologs of FLOWERING LOCUS T are involved in the control of photoperiodic flowering in soybean
- PMID: 20864544
- PMCID: PMC2971601
- DOI: 10.1104/pp.110.160796
Two coordinately regulated homologs of FLOWERING LOCUS T are involved in the control of photoperiodic flowering in soybean
Abstract
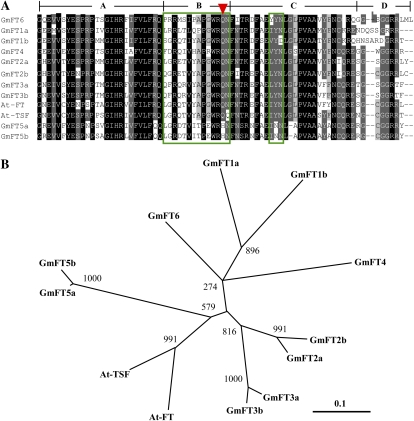
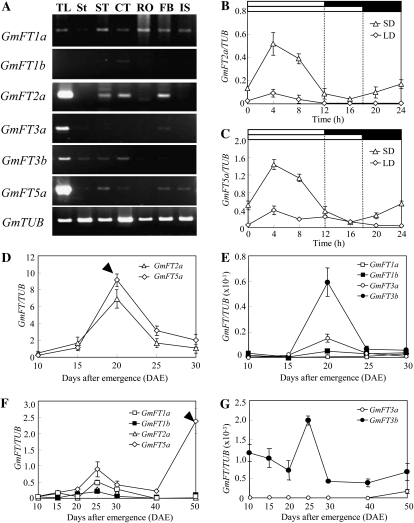
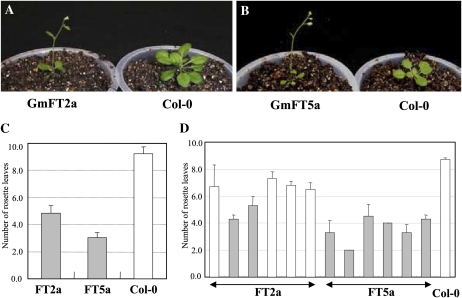
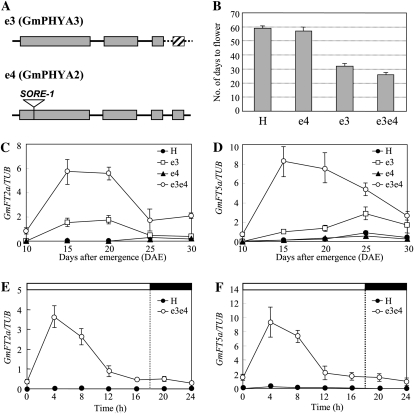
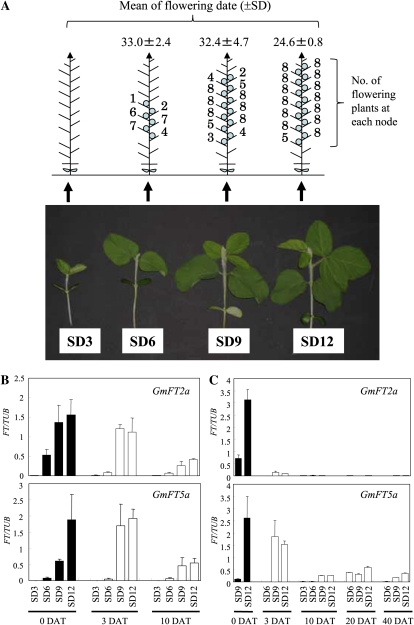
FLOWERING LOCUS T (FT) is a key flowering integrator in Arabidopsis (Arabidopsis thaliana), with homologs that encode florigens in many plant species regardless of the type of photoperiodic response. We identified 10 FT homologs, which were arranged as five pairs of linked genes in different homoeologous chromosomal regions, in soybean (Glycine max), a paleopolyploid species. Two of the FT homologs, GmFT2a and GmFT5a, were highly up-regulated under short-day (SD) conditions (inductive for flowering in soybean) and had diurnal expression patterns with the highest expression 4 h after dawn. Under long-day (LD) conditions, expression of GmFT2a and GmFT5a was down-regulated and did not follow a diurnal pattern. Flowering took much longer to initiate under LD than under SD, and only the GmFT5a transcript accumulated late in development under LD. Ectopic expression analysis in Arabidopsis confirmed that both GmFT2a and GmFT5a had the same function as Arabidopsis FT, but the effect of GmFT5a was more prominent. A double-mutant soybean line for two PHYTOCHROME A (PHYA) genes expressed high levels of GmFT2a and GmFT5a under LD, and it flowered slightly earlier under LD than the wild type grown under SD. The expression levels of GmFT2a and GmFT5a were regulated by the PHYA-mediated photoperiodic regulation system, and the GmFT5a expression was also regulated by a photoperiod-independent system in LD. Taken together, our results suggest that GmFT2a and GmFT5a coordinately control flowering and enable the adaptation of soybean to a wide range of photoperiodic environments.
Figures
References
-
- Abe M, Kobayashi Y, Yamamoto S, Daimon Y, Yamaguchi A, Ikeda Y, Ichinoki H, Notaguchi M, Goto K, Araki T. (2005) FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science 309: 1052–1056 - PubMed
-
- Böhlenius H, Huang T, Charbonnel-Campaa L, Brunner AM, Jansson S, Strauss SH, Nilsson O. (2006) CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 312: 1040–1043 - PubMed
-
- Borthwick HA, Parker MW. (1938) Effectiveness of photoperiodic treatments of plants of different age. Bot Gaz 100: 245–249
-
- Buzzell RI. (1971) Inheritance of a soybean flowering response to fluorescent-daylength conditions. Can J Genet Cytol 13: 703–707
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
