

Evolution under fluctuating environments explains observed robustness in metabolic networks
- PMID: 20865149
- PMCID: PMC2928748
- DOI: 10.1371/journal.pcbi.1000907
Evolution under fluctuating environments explains observed robustness in metabolic networks
Abstract
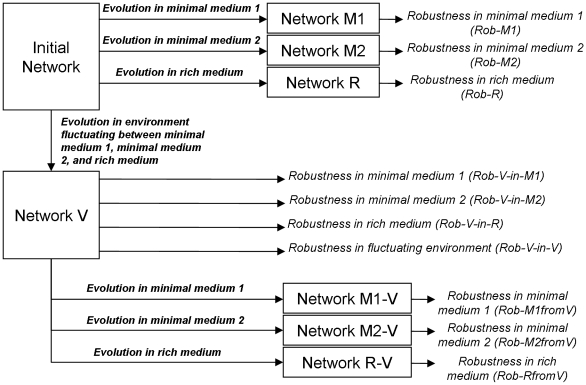
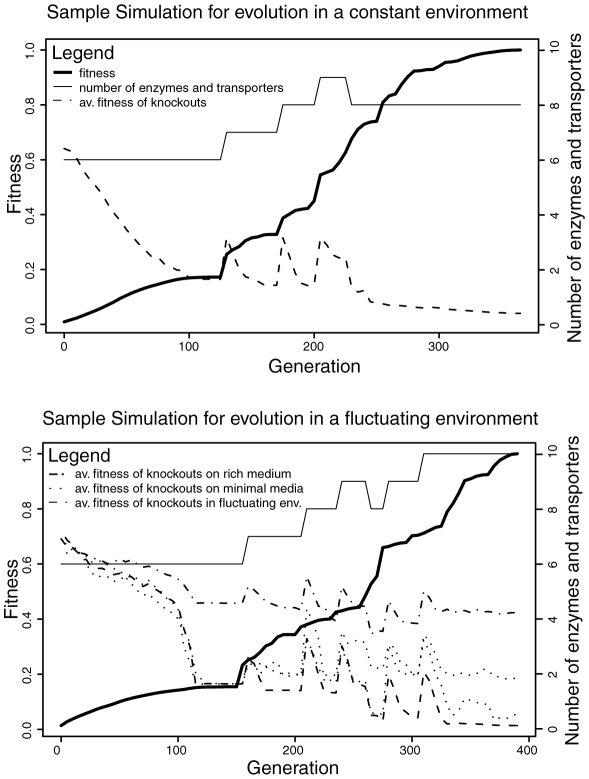
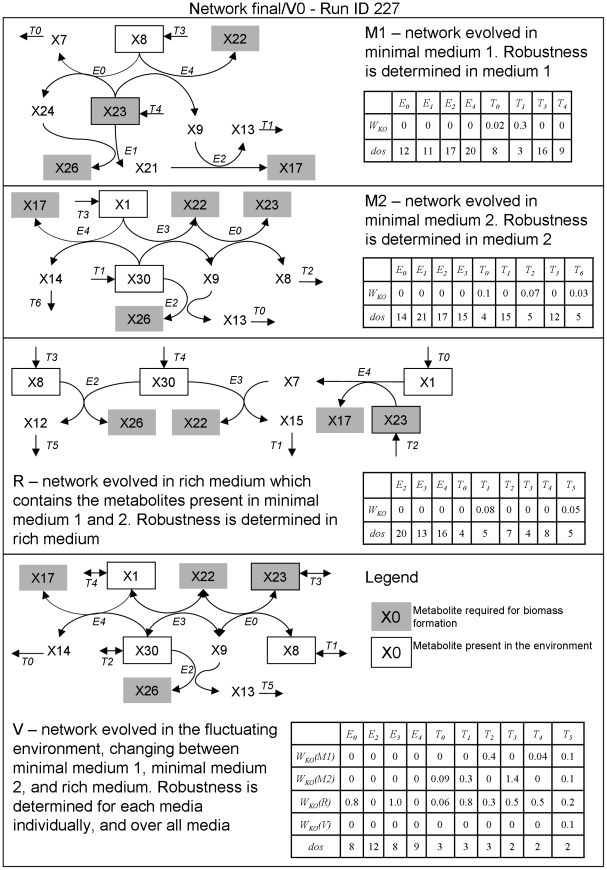
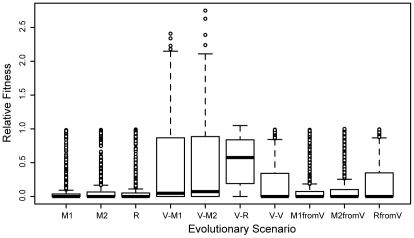
A high level of robustness against gene deletion is observed in many organisms. However, it is still not clear which biochemical features underline this robustness and how these are acquired during evolution. One hypothesis, specific to metabolic networks, is that robustness emerges as a byproduct of selection for biomass production in different environments. To test this hypothesis we performed evolutionary simulations of metabolic networks under stable and fluctuating environments. We find that networks evolved under the latter scenario can better tolerate single gene deletion in specific environments. Such robustness is underlined by an increased number of independent fluxes and multifunctional enzymes in the evolved networks. Observed robustness in networks evolved under fluctuating environments was "apparent," in the sense that it decreased significantly as we tested effects of gene deletions under all environments experienced during evolution. Furthermore, when we continued evolution of these networks under a stable environment, we found that any robustness they had acquired was completely lost. These findings provide evidence that evolution under fluctuating environments can account for the observed robustness in metabolic networks. Further, they suggest that organisms living under stable environments should display lower robustness in their metabolic networks, and that robustness should decrease upon switching to more stable environments.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, et al. Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science. 2003;301:653–657. - PubMed
-
- Giaever G, Chu AM, Ni L, Connelly C, Riles L, et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature. 2002;418:387–391. - PubMed
-
- Kamath RS, Fraser AG, Dong Y, Poulin G, Durbin R, et al. Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature. 2003;421:231–237. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
