

Review
doi: 10.1016/j.cub.2010.08.010.
Building the centriole
Affiliations
- PMID: 20869612
- PMCID: PMC2956124
- DOI: 10.1016/j.cub.2010.08.010
Item in Clipboard
Review
Building the centriole
Curr Biol.
.
Abstract
Centrioles are conserved microtubule-based organelles that lie at the core of the animal centrosome and play a crucial role in nucleating the formation of cilia and flagella in most eukaryotes. Centrioles have a complex ultrastructure with ninefold symmetry and a well-defined length. This structure is assembled from a host of proteins, including a variety of disease gene products. Over a century after the discovery of centrioles, the mechanisms underlying the assembly of these fascinating organelles, in particular the establishment of ninefold symmetry and the control of centriole length, are now starting to be uncovered.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
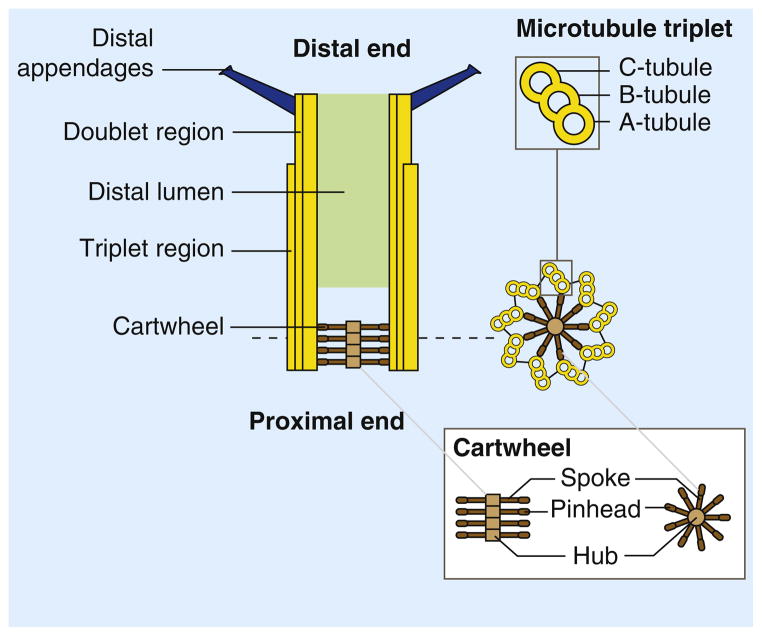
Centrioles are microtubule arrays composed of nine triplets of microtubules organized around a cartwheel structure. The triplets are connected to the cartwheel through the A-tubule, the first to assemble during centriole assembly and the only complete microtubule in a triplet. The B- and C-tubules are incomplete microtubules. In vertebrates and in Chlamydomonas, the C-tubule is shorter than the A- and B-tubules and the distal end of the centriole is thus formed by doublet microtubules [112,129]. The cartwheel is formed by a central hub from which emanate spokes terminated by a pinhead structure that binds the A-tubule of the microtubule triplet. The very distal end of the centriole is decorated by ninefold symmetric distal appendages (or transition fibers) required for anchoring the centrioles at the plasma membrane when they act as a basal bodies.
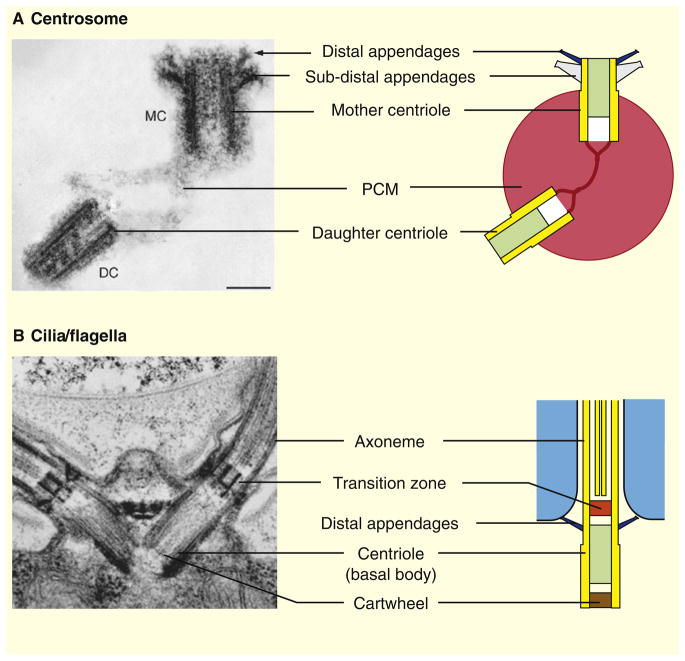
(A) In animal cells, centrioles form the core structure of the centrosome, the main microtubule-organizing center. Quiescent cells (G0) or proliferating cells in the G1 phase of the cell cycle contain a single centrosome. The centrosome is formed by one mature centriole, the mother centriole (MC), and one non-mature centriole, the daughter centriole (DC), linked together and surrounded by a protein matrix called the pericentriolar material (PCM). In vertebrates, the mother centriole is decorated by two sets of ninefold symmetrical appendages: the distal and sub-distal appendages required for ciliogenesis and for the stable anchoring of microtubules at the centrosome, respectively. The distal appendages are conserved throughout eukaryotes, whereas the sub-distal appendages are found only in animal centrosomes. (B) In animals as well as in most other eukaryotes, centrioles are also required for the assembly of cilia/flagella. Centrioles, often referred to as basal bodies in this case, dock to the plasma membrane through their distal appendages and template the assembly of the nine outer microtubule doublets of the axoneme, the cytoskeletal core of cilia/flagella. A distinct structure called the transition zone separates the basal body from the axoneme. Shown are electron micrographs of (A) a human centrosome [130] and (B) the Chlamydomonas flagellar apparatus [24].
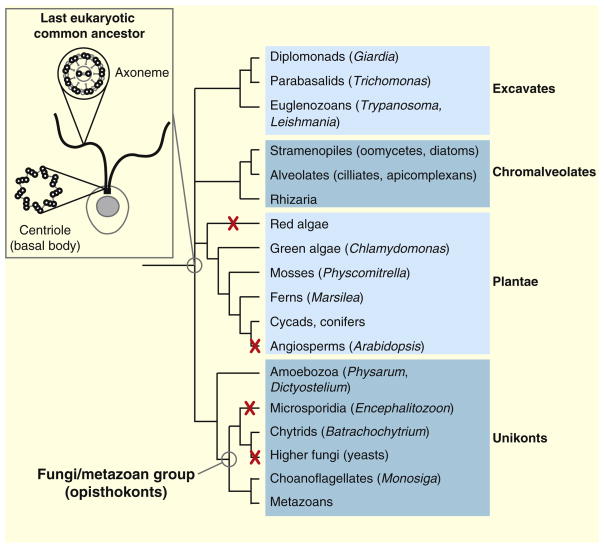
The centriole and the axoneme, i.e. the microtubule core of cilia and flagella, are conserved features of eukaryotes. These structures were probably present in the last eukaryotic common ancestor and are still found in most branches of the eukaryotic tree of life. Centrioles and axonemes were lost concomitantly during evolution of certain taxa, most notably angiosperms and higher fungi. Taxa in which all species have lost centrioles and axonemes are indicated by a red cross. Some taxa, such as amoebozoa, comprise species that form flagella (like Physarum) as well as species completely devoid of centrioles and axonemes (like Dictyostelium). The phylogenetic tree of life is adapted from the Tree of Life web project (http://tolweb.org/tree ). The last eukaryotic common ancestor (inset) is schematically represented as a single-celled organism (the plasma membrane and the nucleus are in gray) bearing two motile flagella (in black). Schematic representations of cross-sections through the centriole and the axoneme are also shown.
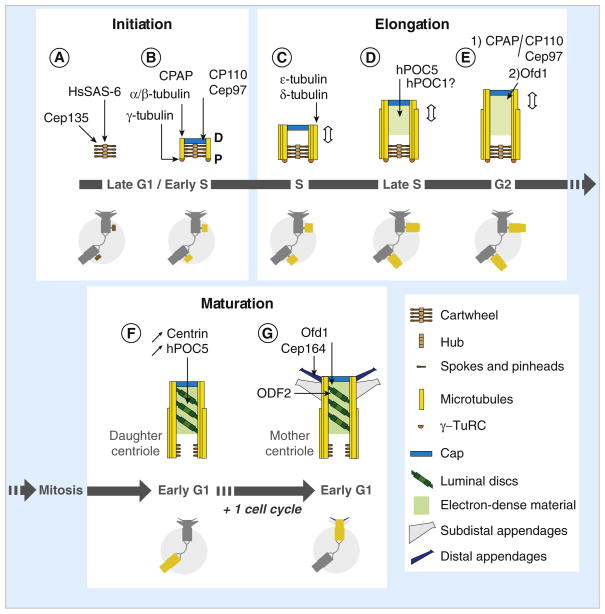
(A) Centriole assembly begins during late G1 or early S phase with the assembly of the cartwheel, which depends on HsSAS-6 for the central hub and Cep135 (Bld10p ortholog) for the spokes and/or the pinheads. (B) CPAP (SAS-4 ortholog) triggers γ-tubulin-dependent nucleation of the A-tubules and their attachment to the pinheads of the cartwheel, possibly by participating in the recruitment of the γ-tubulin ring complex (γ-TuRC) at the proximal end of the cartwheel. Each A-tubule is nucleated by a γ-TuRC and grows unidirectionally from proximal (P) to distal (D). The A-tubule remains capped by the γ-TuRC throughout the assembly process but is lost from daughter and mother centrioles. A cap structure containing CP110 and Cep97 forms at the distal end of the procentriole. The cap is required to control procentriole microtubule growth and probably also to stabilize the nascent procentriole. (C) The B- and C-tubules form by a γ-TuRC-independent mechanism and grow bidirectionally until they reach the length of the A-tubule. The microtubule triplets are stabilized by ε- and δ-tubulin and centriole elongation begins. (D) During S phase, procentrioles elongate up to ~70% of their final length. This step is dependent on hPOC5, and possibly involves hPOC1 as well. (E) Procentriole elongation continues after the transition into G2. Two mechanisms control centriole length at this stage: 1) a balance between the activities of CPAP, which promotes α/β-tubulin incorporation at the distal end, and the cap structure containing CP110 and Cep97; 2) an Ofd1-dependent mechanism. (F) After mitosis, the procentrioles become daughter centrioles. Tilted discs surrounded by electron-dense material are observed from this stage onwards in the distal part of the centriole. Markers like centrin and hPOC5 accumulate within the distal lumen as cell cycle progresses, which could reflect the progressive maturation of the centriole. HsSAS-6 is no longer associated with the proximal part of the daughter centrioles, possibly correlating with the disassembly of the central hub of the cartwheel. In contrast, the spokes could be conserved to some extent as Cep135 remains associated with mother and daughter centrioles. (G) After the second mitosis, centriole maturation is completed when the distal and sub-distal appendages assemble. The assembly of both types of appendage depends on ODF2, a maturation-specific marker recruited at the distal end of the daughter centriole during the previous G2 phase. Assembly of the distal appendages is also dependent on Ofd1, and possibly as well on the distal appendage component Cep164. Schematic representations of the centrosome during the successive steps of centriole assembly and maturation are shown in the lower parts of panels A–G, in which the assembling centrioles are highlighted (brown when only the cartwheel is present, yellow for later stages).
References
-
- Gönczy P, Echeverri C, Oegema K, Coulson A, Jones SJ, Copley RR, Duperon J, Oegema J, Brehm M, Cassin E, et al. Functional genomic analysis of cell division in C. elegans using RNAi of genes on chromosome III. Nature. 2000;408:331–336. - PubMed
-
- Kirkham M, Muller-Reichert T, Oegema K, Grill S, Hyman A. SAS-4 is a C. elegans centriolar protein that controls centrosome size. Cell. 2003;112:575–587. - PubMed
-
- Leidel S, Gönczy P. SAS-4 is essential for centrosome duplication in C. elegans and is recruited to daughter centrioles once per cell cycle. Dev Cell. 2003;4:431–439. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
