

Forcing form and function: biomechanical regulation of tumor evolution
- PMID: 20870407
- PMCID: PMC3014395
- DOI: 10.1016/j.tcb.2010.08.015
Forcing form and function: biomechanical regulation of tumor evolution
Abstract
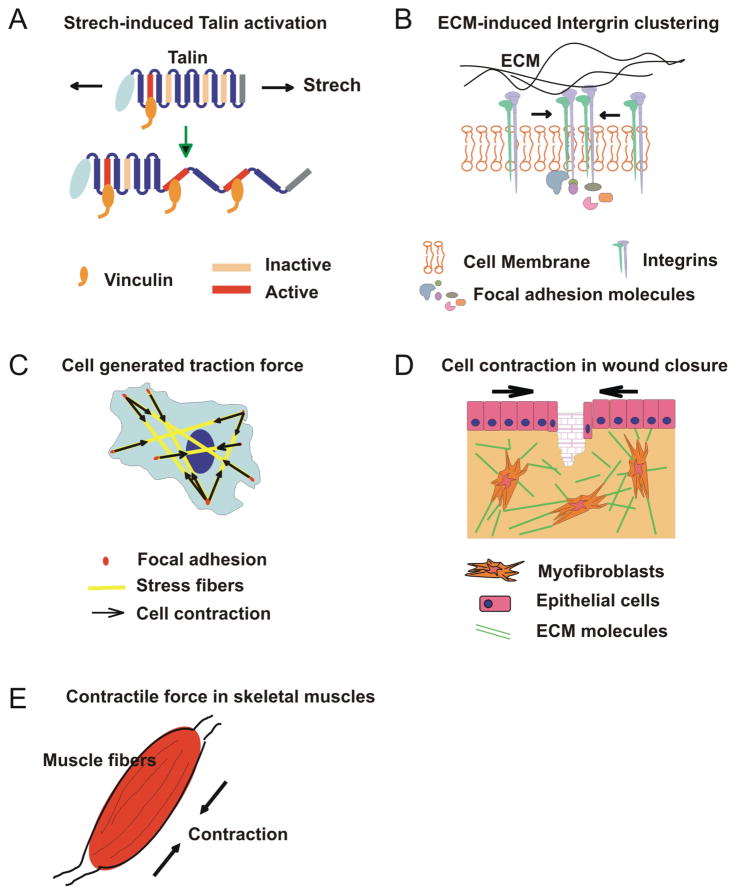
Cancer cells exist in a constantly evolving tissue microenvironment of diverse cell types within a proteinaceous extracellular matrix. As tumors evolve, the physical forces within this complex microenvironment change, with pleiotropic effects on both cell- and tissue-level behaviors. Recent work suggests that these biomechanical factors direct tissue development and modulate tissue homeostasis, and, when altered, crucially influence tumor evolution. In this review, we discuss the biomechanical regulation of cell and tissue homeostasis from the molecular, cellular and tissue levels, including how modifications of this physical dialogue could contribute to cancer etiology. Because of the broad impact of biomechanical factors on cell and tissue functions, an understanding of tumor evolution from the biomechanical perspective should improve risk assessment, clinical diagnosis and the efficacy of cancer treatment.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
