

Substrates of the Arabidopsis thaliana protein isoaspartyl methyltransferase 1 identified using phage display and biopanning
- PMID: 20870712
- PMCID: PMC2988334
- DOI: 10.1074/jbc.M110.157008
Substrates of the Arabidopsis thaliana protein isoaspartyl methyltransferase 1 identified using phage display and biopanning
Abstract
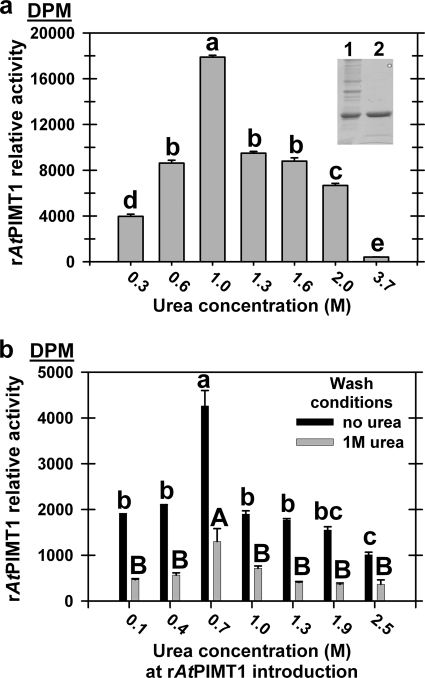
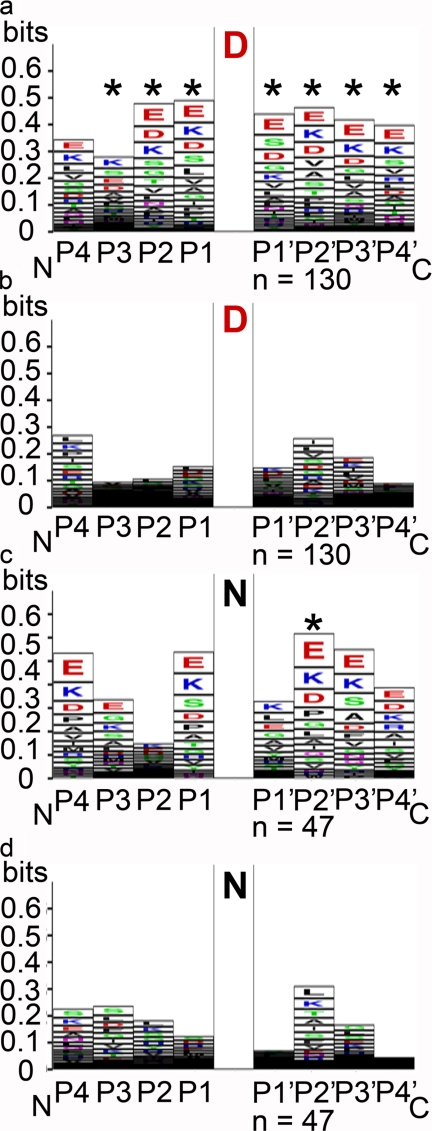
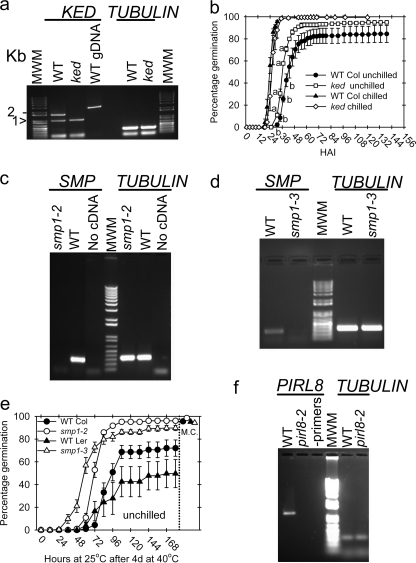
The role of protein isoaspartyl methyltransferase (PIMT) in repairing a wide assortment of damaged proteins in a host of organisms has been inferred from the affinity of the enzyme for isoaspartyl residues in a plethora of amino acid contexts. The identification of PIMT target proteins in plant seeds, where the enzyme is highly active and proteome long-lived, has been hindered by large amounts of isoaspartate-containing storage proteins. Mature seed phage display libraries circumvented this problem. Inclusion of the PIMT co-substrate, S-adenosylmethionine (AdoMet), during panning permitted PIMT to retain aged phage in greater numbers than controls lacking co-substrate or when PIMT protein binding was poisoned with S-adenosyl homocysteine. After four rounds, phage titer plateaued in AdoMet-containing pans, whereas titer declined in both controls. This strategy identified 17 in-frame PIMT target proteins, including a cupin-family protein similar to those identified previously using on-blot methylation. All recovered phage had at least one susceptible Asp or Asn residue. Five targets were recovered independently. Two in-frame targets were produced in Escherichia coli as recombinant proteins and shown by on-blot methylation to acquire isoAsp, becoming a PIMT target. Both gained isoAsp rapidly in solution upon thermal insult. Mutant analysis of plants deficient in any of three in-frame PIMT targets resulted in demonstrable phenotypes. An over-representation of clones encoding proteins involved in protein production suggests that the translational apparatus comprises a subgroup for which PIMT-mediated repair is vital for orthodox seed longevity. Impaired PIMT activity would hinder protein function in these targets, possibly resulting in poor seed performance.
Figures



Similar articles
-
Arabidopsis protein l-ISOASPARTYL METHYLTRANSFERASE repairs isoaspartyl damage to antioxidant enzymes and increases heat and oxidative stress tolerance.J Biol Chem. 2020 Jan 17;295(3):783-799. doi: 10.1074/jbc.RA119.010779. Epub 2019 Dec 12. J Biol Chem. 2020. PMID: 31831624 Free PMC article.
-
An Arabidopsis ATP-dependent, DEAD-box RNA helicase loses activity upon IsoAsp formation but is restored by PROTEIN ISOASPARTYL METHYLTRANSFERASE.Plant Cell. 2013 Jul;25(7):2573-86. doi: 10.1105/tpc.113.113456. Epub 2013 Jul 31. Plant Cell. 2013. PMID: 23903319 Free PMC article.
-
Protein repair L-isoaspartyl methyltransferase 1 is involved in both seed longevity and germination vigor in Arabidopsis.Plant Cell. 2008 Nov;20(11):3022-37. doi: 10.1105/tpc.108.058479. Epub 2008 Nov 14. Plant Cell. 2008. PMID: 19011119 Free PMC article.
-
PROTEIN l-ISOASPARTYL METHYLTRANSFERASE (PIMT) in plants: regulations and functions.Biochem J. 2020 Nov 27;477(22):4453-4471. doi: 10.1042/BCJ20200794. Biochem J. 2020. PMID: 33245750 Review.
-
PIMT-Mediated Protein Repair: Mechanism and Implications.Biochemistry (Mosc). 2019 May;84(5):453-463. doi: 10.1134/S0006297919050018. Biochemistry (Mosc). 2019. PMID: 31234761 Review.
Cited by
-
Uses of phage display in agriculture: a review of food-related protein-protein interactions discovered by biopanning over diverse baits.Comput Math Methods Med. 2013;2013:653759. doi: 10.1155/2013/653759. Epub 2013 Apr 28. Comput Math Methods Med. 2013. PMID: 23710253 Free PMC article. Review.
-
Arabidopsis PIRL6 Is Essential for Male and Female Gametogenesis and Is Regulated by Alternative Splicing.Plant Physiol. 2018 Nov;178(3):1154-1169. doi: 10.1104/pp.18.00329. Epub 2018 Sep 11. Plant Physiol. 2018. PMID: 30206104 Free PMC article.
-
PROTEIN L-ISOASPARTYL METHYLTRANSFERASE2 is differentially expressed in chickpea and enhances seed vigor and longevity by reducing abnormal isoaspartyl accumulation predominantly in seed nuclear proteins.Plant Physiol. 2013 Mar;161(3):1141-57. doi: 10.1104/pp.112.206243. Epub 2013 Jan 2. Plant Physiol. 2013. PMID: 23284083 Free PMC article.
-
Late Embryogenesis Abundant Protein-Client Protein Interactions.Plants (Basel). 2020 Jun 29;9(7):814. doi: 10.3390/plants9070814. Plants (Basel). 2020. PMID: 32610443 Free PMC article. Review.
-
Comparison of Protein N-Homocysteinylation in Rat Plasma under Elevated Homocysteine Using a Specific Chemical Labeling Method.Molecules. 2016 Sep 8;21(9):1195. doi: 10.3390/molecules21091195. Molecules. 2016. PMID: 27617989 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
