

Identification of a lysosomal pathway regulating degradation of the bone morphogenetic protein receptor type II
- PMID: 20870717
- PMCID: PMC2988369
- DOI: 10.1074/jbc.M110.132415
Identification of a lysosomal pathway regulating degradation of the bone morphogenetic protein receptor type II
Abstract
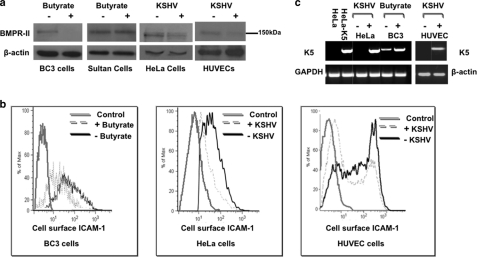
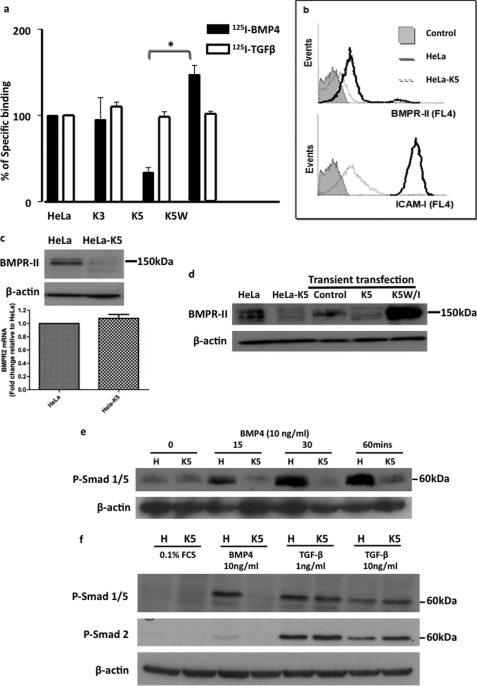
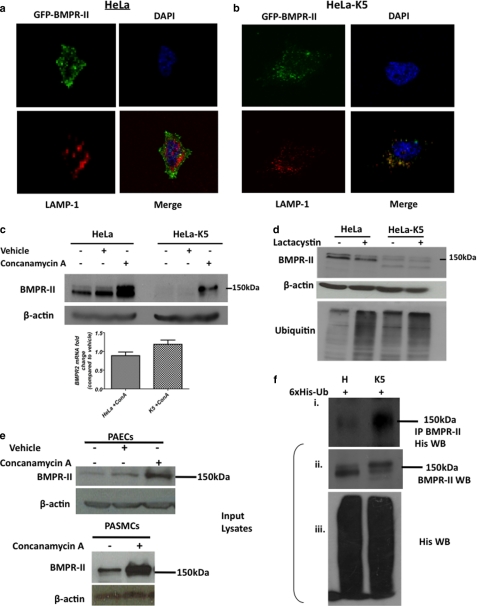
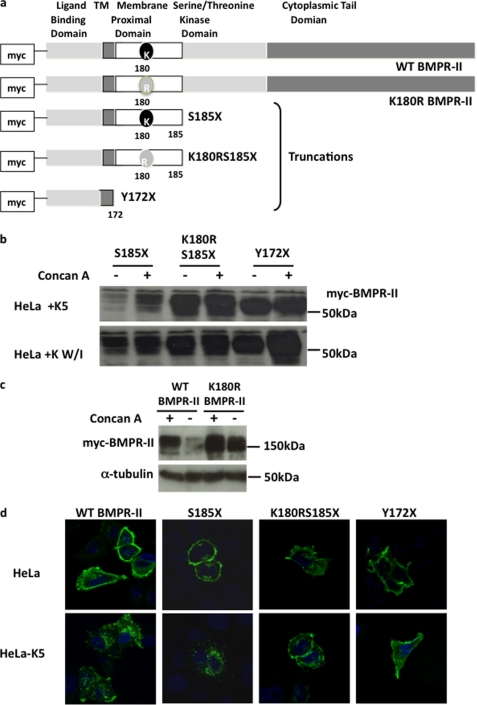
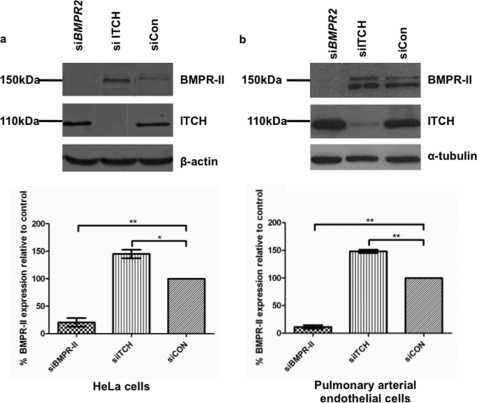
Bone morphogenetic proteins (BMPs) are critically involved in early development and cell differentiation. In humans, dysfunction of the bone morphogenetic protein type II receptor (BMPR-II) is associated with pulmonary arterial hypertension (PAH) and neoplasia. The ability of Kaposi sarcoma-associated herpesvirus (KSHV), the etiologic agent of Kaposi sarcoma and primary effusion lymphoma, to down-regulate cell surface receptor expression is well documented. Here we show that KSHV infection reduces cell surface BMPR-II. We propose that this occurs through the expression of the viral lytic gene, K5, a ubiquitin E3 ligase. Ectopic expression of K5 leads to BMPR-II ubiquitination and lysosomal degradation with a consequent decrease in BMP signaling. The down-regulation by K5 is dependent on both its RING domain and a membrane-proximal lysine in the cytoplasmic domain of BMPR-II. We demonstrate that expression of BMPR-II protein is constitutively regulated by lysosomal degradation in vascular cells and provide preliminary evidence for the involvement of the mammalian E3 ligase, Itch, in the constitutive degradation of BMPR-II. Disruption of BMP signaling may therefore play a role in the pathobiology of diseases caused by KSHV infection, as well as KSHV-associated tumorigenesis and vascular disease.
Figures
References
-
- Miyazono K., Maeda S., Imamura T. (2005) Cytokine Growth Factor Rev. 16, 251–263 - PubMed
-
- Beppu H., Kawabata M., Hamamoto T., Chytil A., Minowa O., Noda T., Miyazono K. (2000) Dev. Biol. 221, 249–258 - PubMed
-
- Liu D., Wang J., Kinzel B., Müeller M., Mao X., Valdez R., Liu Y., Li E. (2007) Blood 110, 1502–1510 - PubMed
-
- International, PPH Consortium Lane K. B., Machado R. D., Pauciulo M. W., Thomson J. R., Philips J. A., 3rd, Loyd J. E., Nichols W. C., Trembath R. C. (2000) Nat. Genet. 26, 81–84 - PubMed
-
- Thomson J. R., Machado R. D., Pauciulo M. W., Morgan N. V., Humbert M., Elliott G. C., Ward K., Yacoub M., Mikhail G., Rogers P., Newman J., Wheeler L., Higenbottam T., Gibbs J. S., Egan J., Crozier A., Peacock A., Allcock R., Corris P., Loyd J. E., Trembath R. C., Nichols W. C. (2000) J. Med. Genet. 37, 741–745 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
