

Emergence of cortical inhibition by coordinated sensory-driven plasticity at distinct synaptic loci
- PMID: 20871602
- PMCID: PMC2950257
- DOI: 10.1038/nn.2639
Emergence of cortical inhibition by coordinated sensory-driven plasticity at distinct synaptic loci
Abstract
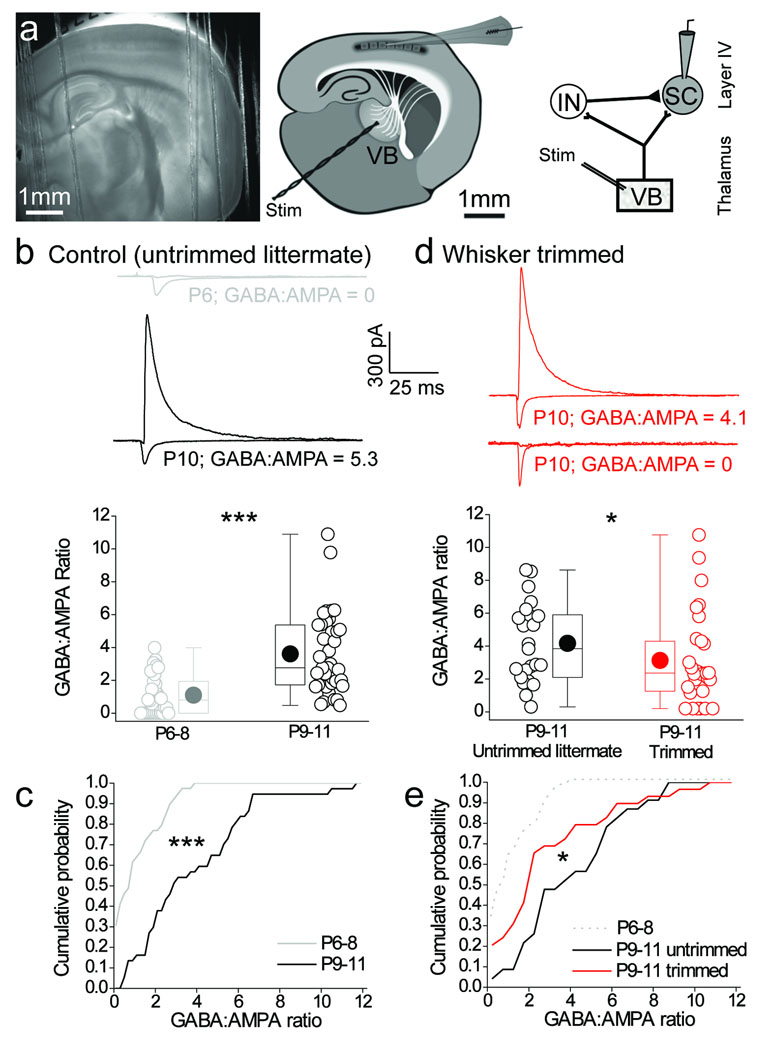
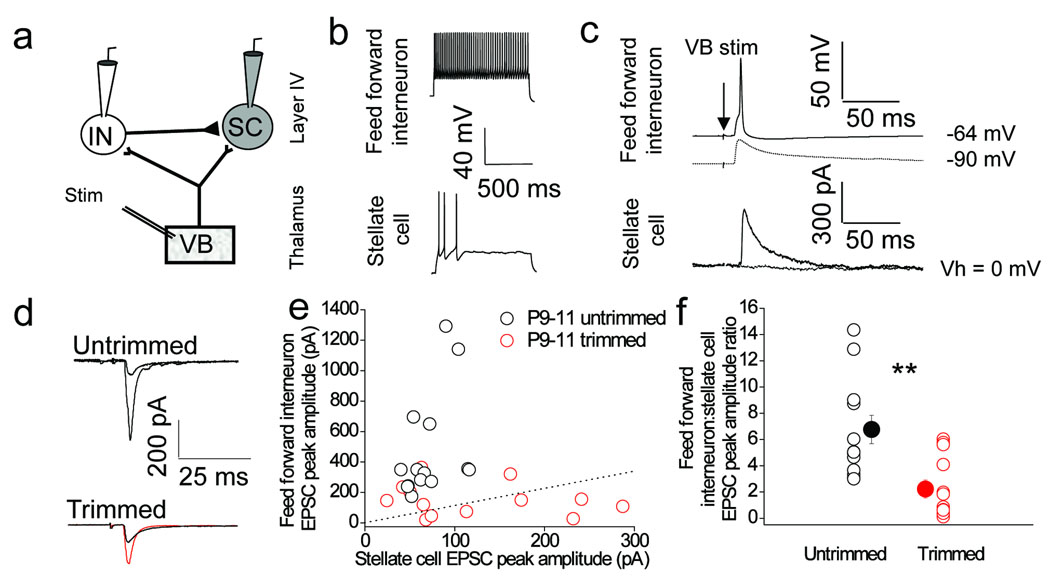
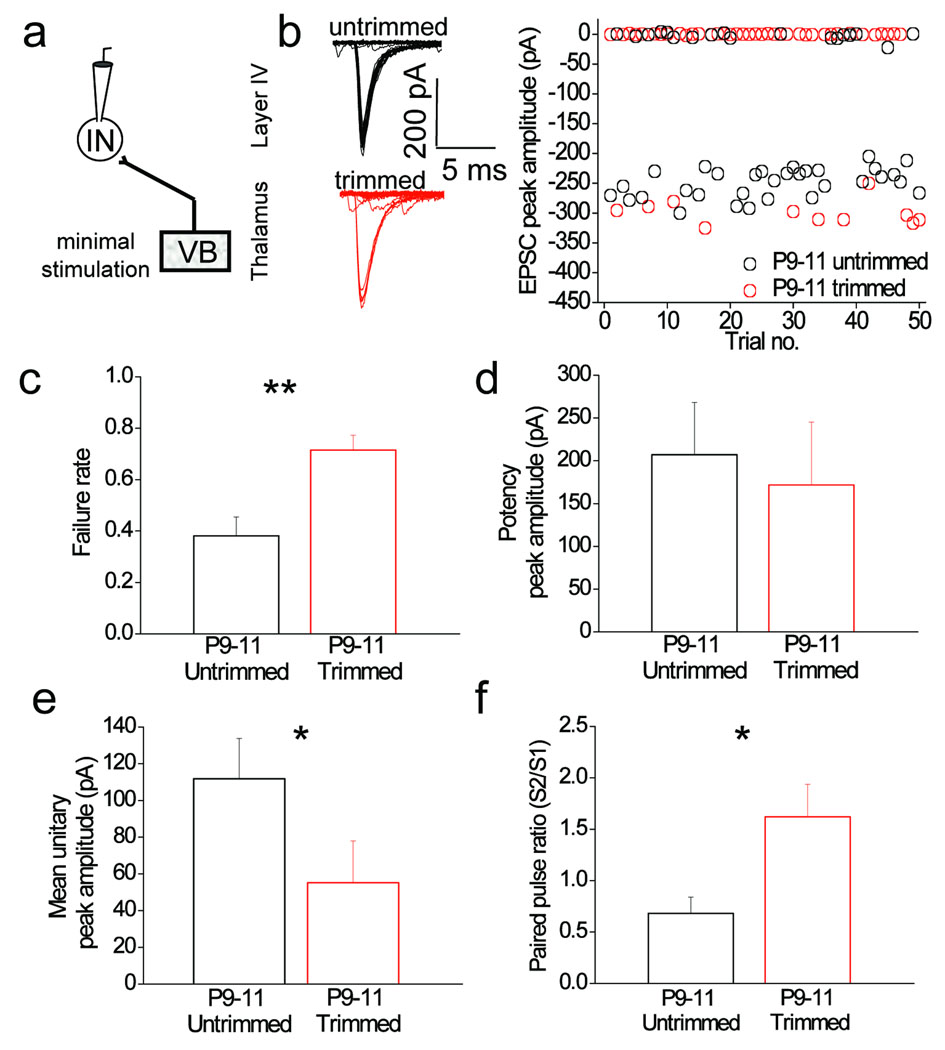
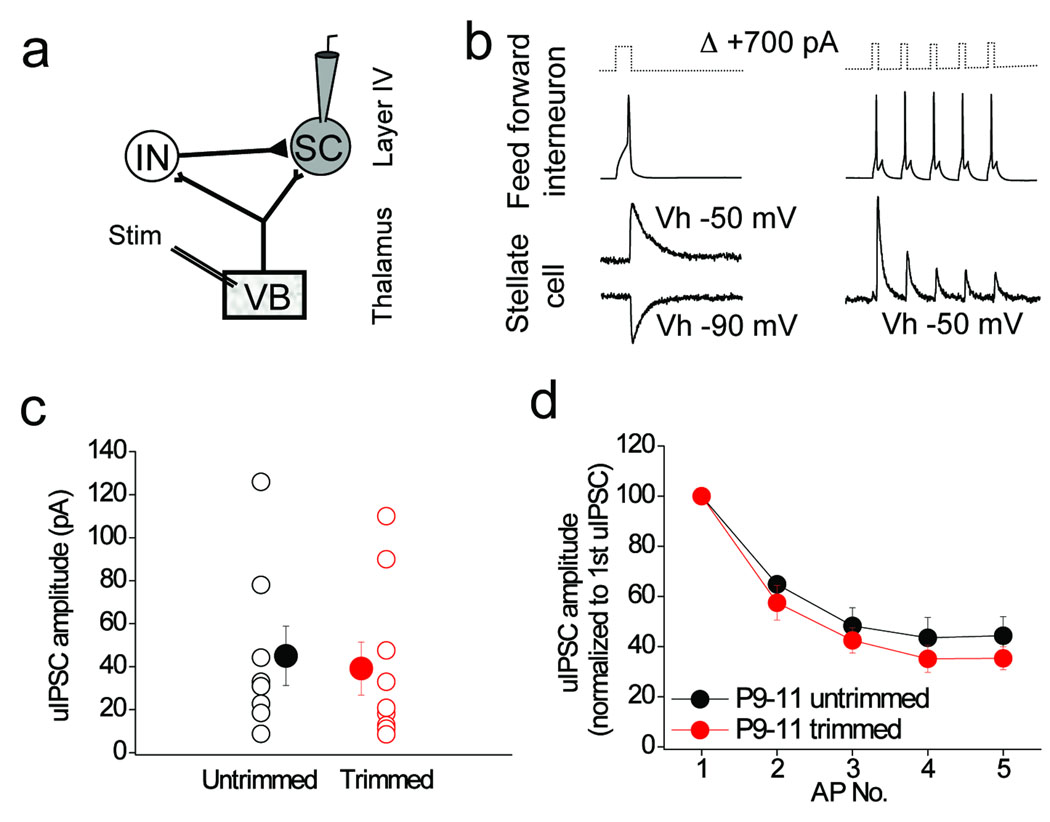
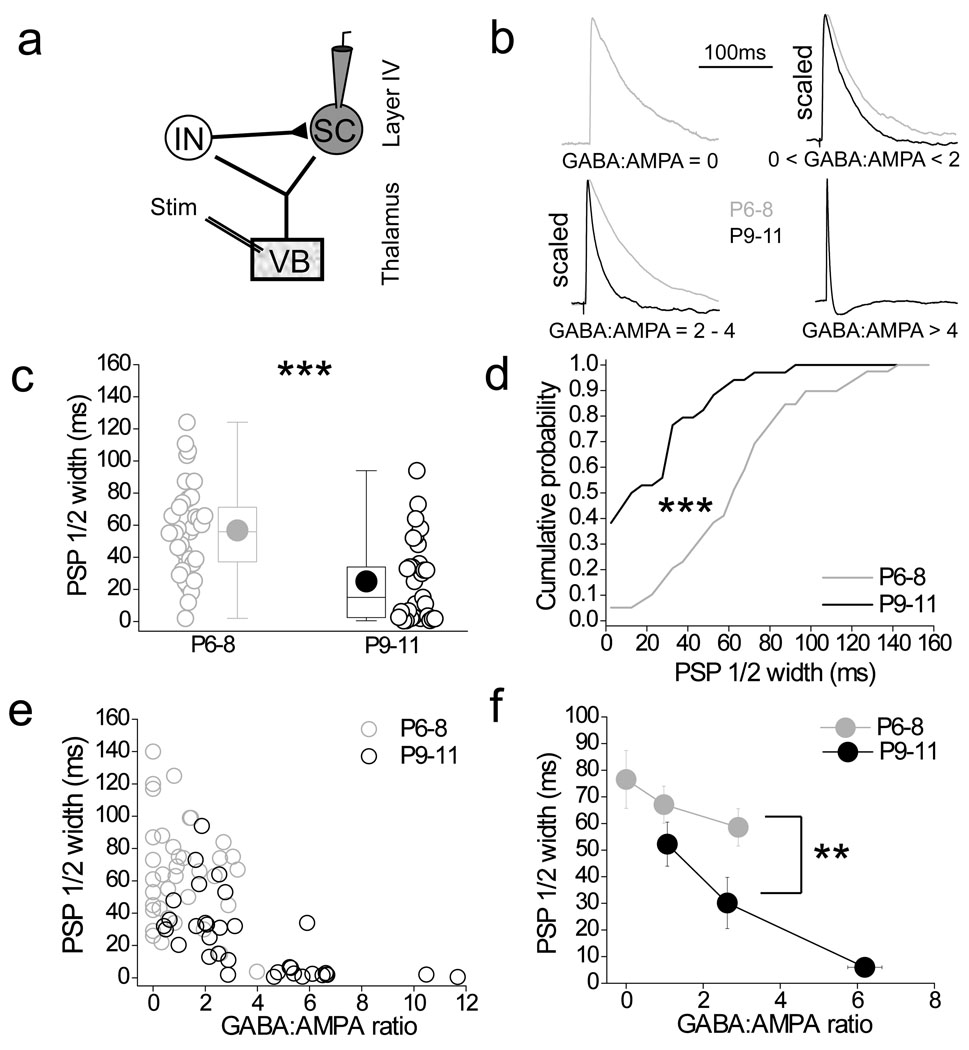
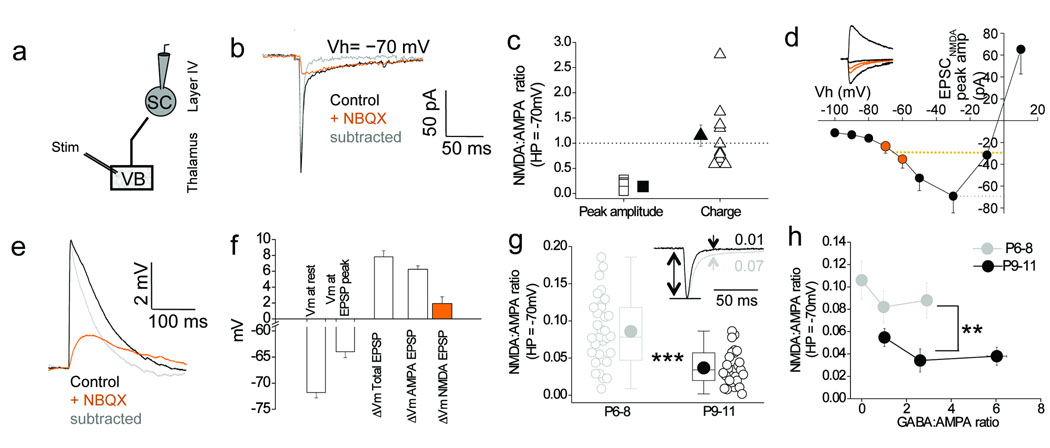
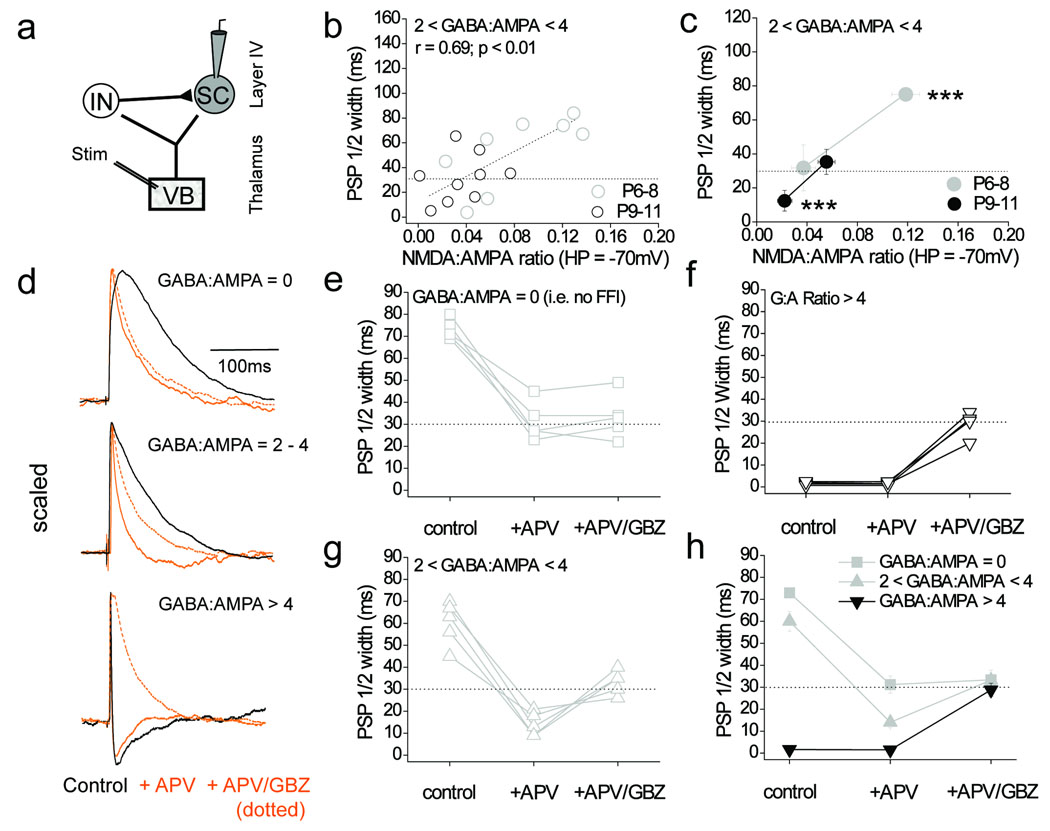
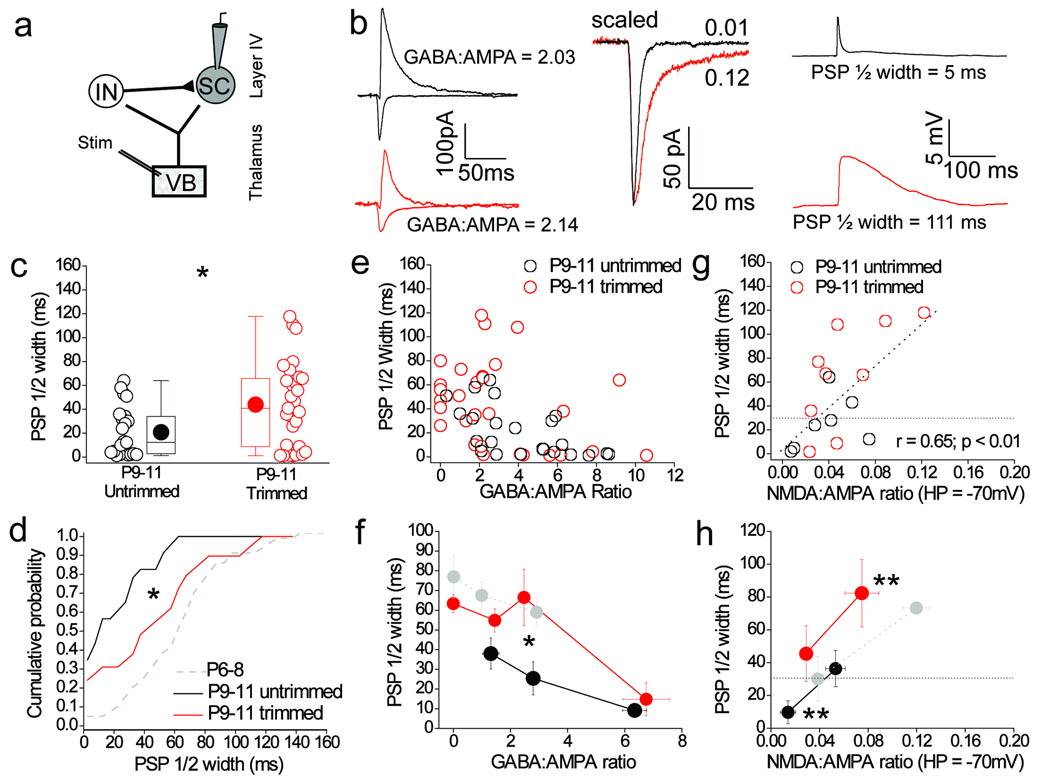
Feedforward GABAergic inhibition sets the dendritic integration window, thereby controlling timing and output in cortical circuits. However, the manner in which feedforward inhibitory circuits emerge is unclear, despite this being a critical step for neocortical development and function. We found that sensory experience drove plasticity of the feedforward inhibitory circuit in mouse layer 4 somatosensory barrel cortex in the second postnatal week via two distinct mechanisms. First, sensory experience selectively strengthened thalamocortical-to-feedforward interneuron inputs via a presynaptic mechanism but did not regulate other inhibitory circuit components. Second, experience drove a postsynaptic mechanism in which a downregulation of a prominent thalamocortical NMDA excitatory postsynaptic potential in stellate cells regulated the final expression of functional feedforward inhibitory input. Thus, experience is required for specific, coordinated changes at thalamocortical synapses onto both inhibitory and excitatory neurons, producing a circuit plasticity that results in maturation of functional feedforward inhibition in layer 4.
Figures
References
-
- Woolsey TA, Van der Loos H. The structural organization of layer IV in the somatosensory region (SI) of mouse cerebral cortex. The description of a cortical field composed of discrete cytoarchitectonic units. Brain Res. 1970;17:205–242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
