

Immunocytochemical determination of the subcellular distribution of ascorbate in plants
- PMID: 20872269
- PMCID: PMC3015205
- DOI: 10.1007/s00425-010-1275-x
Immunocytochemical determination of the subcellular distribution of ascorbate in plants
Abstract
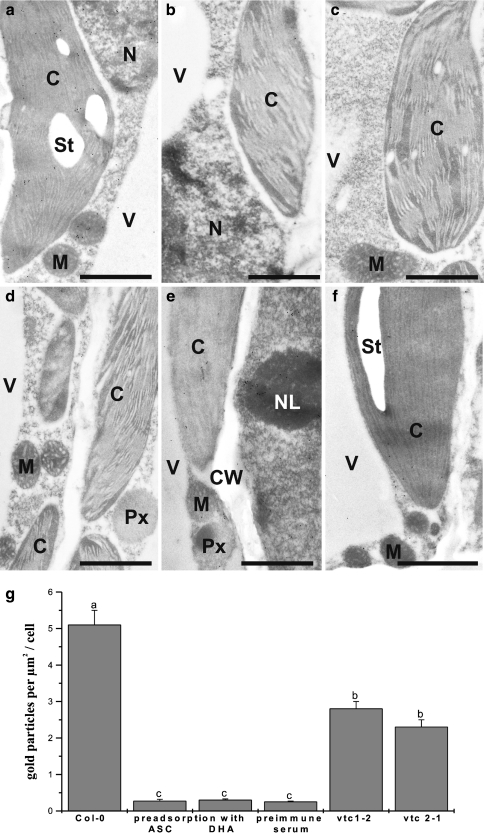
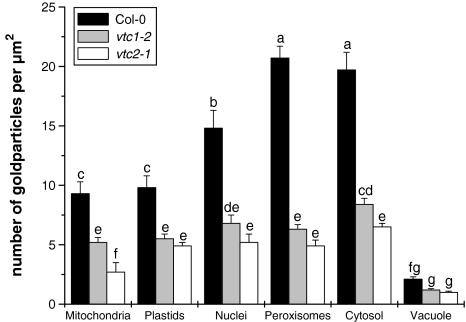
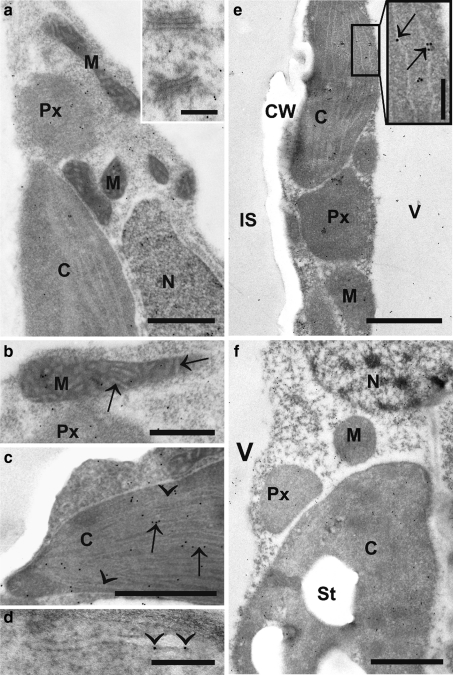
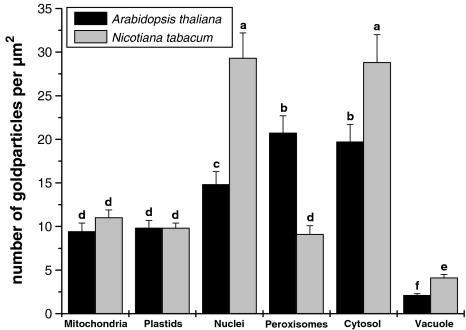
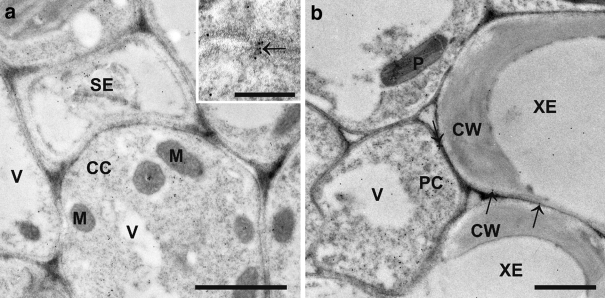
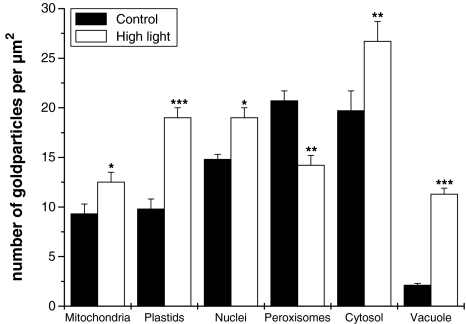
Ascorbate is an important antioxidant in plants and fulfills many functions related to plant defense, redox signaling and modulation of gene expression. We have analyzed the subcellular distribution of reduced and oxidized ascorbate in leaf cells of Arabidopsis thaliana and Nicotiana tabacum by high-resolution immuno electron microscopy. The accuracy and specificity of the applied method is supported by several observations. First, preadsorption of the ascorbate antisera with ascorbic acid or dehydroascorbic acid resulted in the reduction of the labeling to background levels. Second, the overall labeling density was reduced between 50 and 61% in the ascorbate-deficient Arabidopsis mutants vtc1-2 and vtc2-1, which correlated well with biochemical measurements. The highest ascorbate-specific labeling was detected in nuclei and the cytosol whereas the lowest levels were found in vacuoles. Intermediate labeling was observed in chloroplasts, mitochondria and peroxisomes. This method was used to determine the subcellular ascorbate distribution in leaf cells of plants exposed to high light intensity, a stress factor that is well known to cause an increase in cellular ascorbate concentration. High light intensities resulted in a strong increase in overall labeling density. Interestingly, the strongest compartment-specific increase was found in vacuoles (fourfold) and in plastids (twofold). Ascorbate-specific labeling was restricted to the matrix of mitochondria and to the stroma of chloroplasts in control plants but was also detected in the lumen of thylakoids after high light exposure. In summary, this study reveals an improved insight into the subcellular distribution of ascorbate in plants and the method can now be applied to determine compartment-specific changes in ascorbate in response to various stress situations.
Figures
References
-
- Baier M, Kandlbinder A, Golldack D, Dietz KJ. Oxidative stress and ozone: perception, signalling and response. Plant Cell Environ. 2005;28:1012–1020. doi: 10.1111/j.1365-3040.2005.01326.x. - DOI
-
- Chamseddine M, Wided BA, Guy H, Marie-Edith C, Fatma J. Cadmium and copper induction of oxidative stress and antioxidative response in tomato (Solanum lycopersicon) leaves. Plant Growth Regul. 2009;27:89–99. doi: 10.1007/s10725-008-9324-1. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
