

Loss of ethanol conditioned taste aversion and motor stimulation in knockin mice with ethanol-insensitive α2-containing GABA(A) receptors
- PMID: 20876231
- PMCID: PMC3014308
- DOI: 10.1124/jpet.110.171645
Loss of ethanol conditioned taste aversion and motor stimulation in knockin mice with ethanol-insensitive α2-containing GABA(A) receptors
Abstract
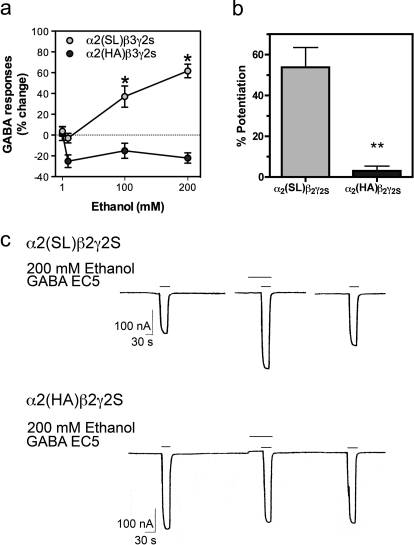
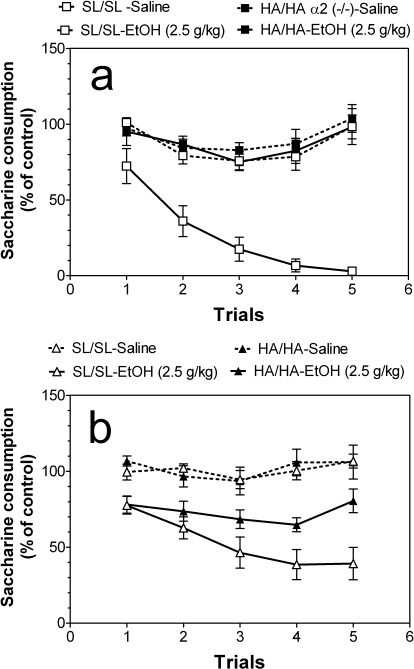
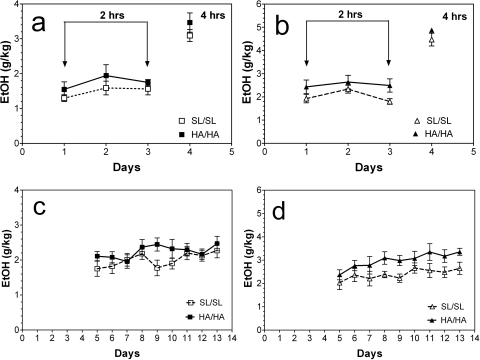
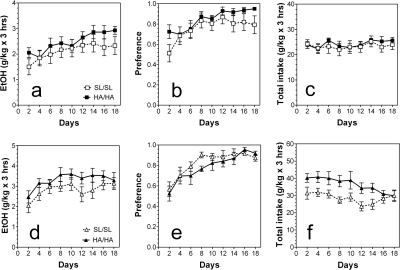
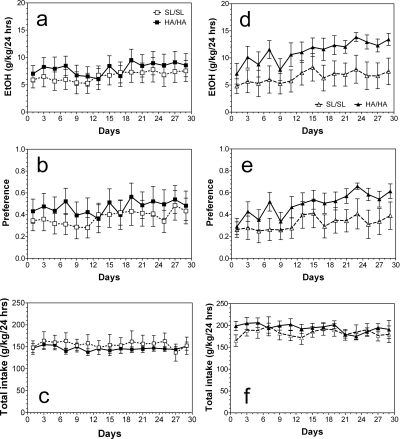
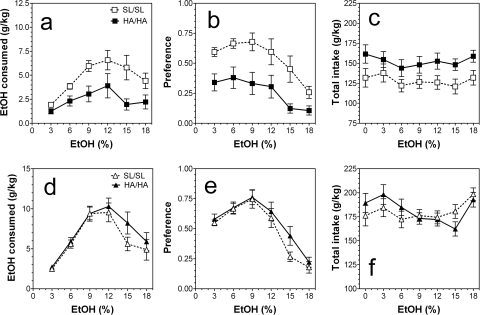
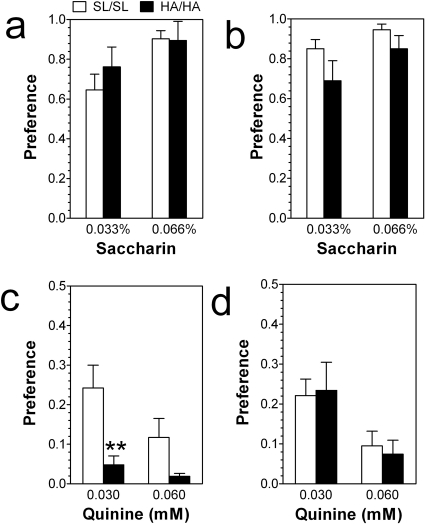
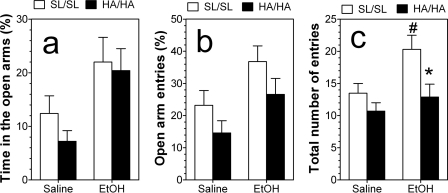
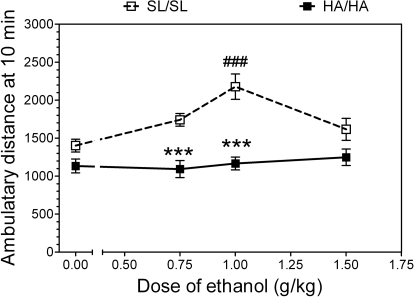
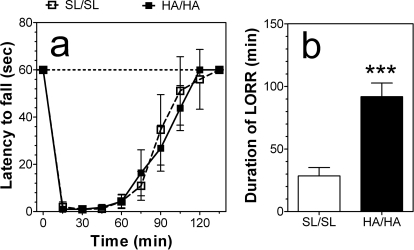
GABA type A receptors (GABA(A)-Rs) are potential targets of ethanol. However, there are multiple subtypes of this receptor, and, thus far, individual subunits have not been definitively linked with specific ethanol behavioral actions. Interestingly, though, a chromosomal cluster of four GABA(A)-R subunit genes, including α2 (Gabra2), was associated with human alcoholism (Am J Hum Genet 74:705-714, 2004; Pharmacol Biochem Behav 90:95-104, 2008; J Psychiatr Res 42:184-191, 2008). The goal of our study was to determine the role of receptors containing this subunit in alcohol action. We designed an α2 subunit with serine 270 to histidine and leucine 277 to alanine mutations that was insensitive to potentiation by ethanol yet retained normal GABA sensitivity in a recombinant expression system. Knockin mice containing this mutant subunit were tested in a range of ethanol behavioral tests. These mutant mice did not develop the typical conditioned taste aversion in response to ethanol and showed complete loss of the motor stimulant effects of ethanol. Conversely, they also demonstrated changes in ethanol intake and preference in multiple tests. The knockin mice showed increased ethanol-induced hypnosis but no difference in anxiolytic effects or recovery from acute ethanol-induced motor incoordination. Overall, these studies demonstrate that the effects of ethanol at GABAergic synapses containing the α2 subunit are important for specific behavioral effects of ethanol that may be relevant to the genetic linkage of this subunit with human alcoholism.
Figures
References
-
- Blednov YA, Walker D, Alva H, Creech K, Findlay G, Harris RA. (2003) GABAA receptor α1 and β2 subunit null mutant mice: behavioral responses to ethanol. J Pharmacol Exp Ther 305:854–863 - PubMed
-
- Boehm SL, 2nd, Ponomarev I, Jennings AW, Whiting PJ, Rosahl TW, Garrett EM, Blednov YA, Harris RA. (2004) γ-Aminobutyric acid A receptor subunit mutant mice: new perspectives on alcohol actions. Biochem Pharmacol 68:1581–1602 - PubMed
-
- Borghese CM, Werner DF, Topf N, Baron NV, Henderson LA, Boehm SL, 2nd, Blednov YA, Saad A, Dai S, Pearce RA, et al. (2006) An isoflurane- and alcohol-insensitive mutant GABA(A) receptor α(1) subunit with near-normal apparent affinity for GABA: characterization in heterologous systems and production of knockin mice. J Pharmacol Exp Ther 319:208–218 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
