

The C. elegans peroxidasin PXN-2 is essential for embryonic morphogenesis and inhibits adult axon regeneration
- PMID: 20876652
- PMCID: PMC2964093
- DOI: 10.1242/dev.049189
The C. elegans peroxidasin PXN-2 is essential for embryonic morphogenesis and inhibits adult axon regeneration
Abstract
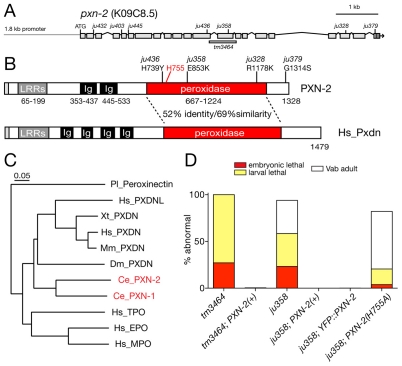
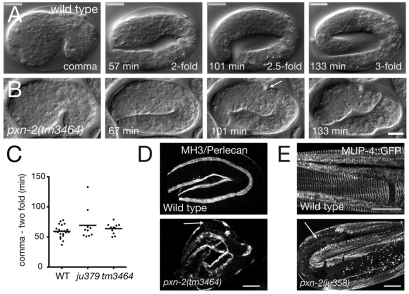
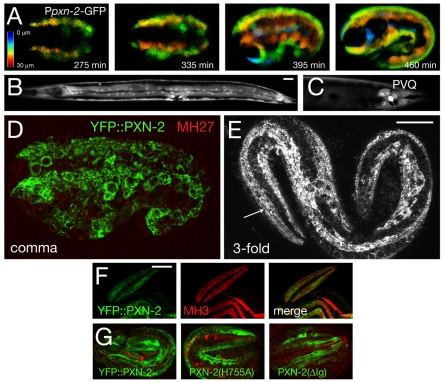
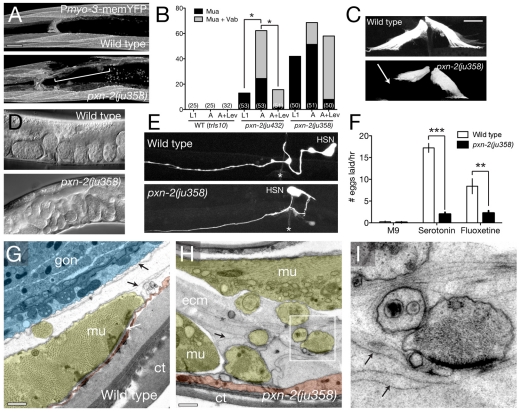
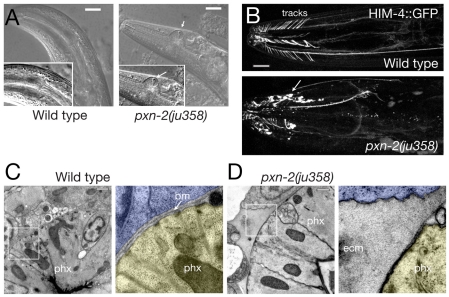
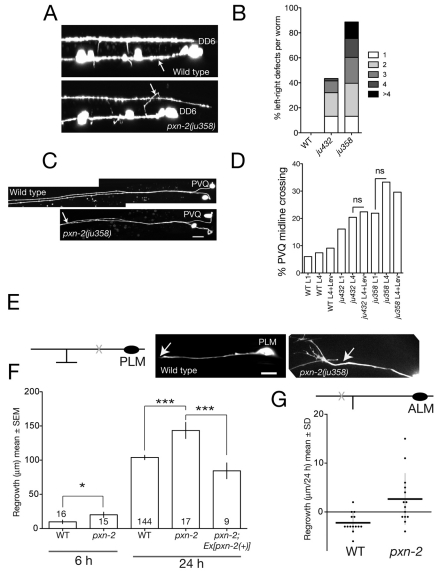
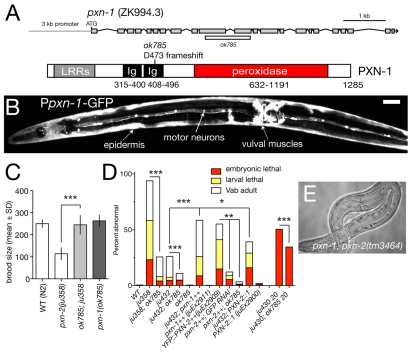
Peroxidasins form a highly conserved family of extracellular peroxidases of unknown cellular function. We identified the C. elegans peroxidasin PXN-2 in screens for mutants defective in embryonic morphogenesis. We find that PXN-2 is essential for specific stages of embryonic morphogenesis and muscle-epidermal attachment, and is also required postembryonically for basement membrane integrity. The peroxidase catalytic activity of PXN-2 is necessary for these developmental roles. pxn-2 mutants display aberrant ultrastructure of the extracellular matrix, suggesting a role in basement membrane consolidation. PXN-2 affects specific axon guidance choice points in the developing nervous system but is dispensable for maintenance of process positions. In adults, loss of pxn-2 function promotes regrowth of axons after injury, providing the first evidence that C. elegans extracellular matrix can play an inhibitory role in axon regeneration. Loss of function in the closely related C. elegans peroxidasin pxn-1 does not cause overt developmental defects. Unexpectedly, pxn-2 mutant phenotypes are suppressed by loss of function in pxn-1 and exacerbated by overexpression of wild-type pxn-1, indicating that PXN-1 and PXN-2 have antagonistic functions. These results demonstrate that peroxidasins play crucial roles in development and reveal a new role for peroxidasins as extracellular inhibitors of axonal regeneration.
Figures
Similar articles
-
pxn-1 and pxn-2 May Interact Negatively during Neuronal Development and Aging in C. elegans.Mol Cells. 2015 Aug;38(8):729-33. doi: 10.14348/molcells.2015.0124. Epub 2015 Jul 21. Mol Cells. 2015. PMID: 26194821 Free PMC article.
-
Genetic Suppression of Basement Membrane Defects in Caenorhabditis elegans by Gain of Function in Extracellular Matrix and Cell-Matrix Attachment Genes.Genetics. 2018 Apr;208(4):1499-1512. doi: 10.1534/genetics.118.300731. Epub 2018 Feb 12. Genetics. 2018. PMID: 29440357 Free PMC article.
-
A role for peroxidasin PXN-1 in aspects of C. elegans development.Mol Cells. 2015 Jan 31;38(1):51-7. doi: 10.14348/molcells.2015.2202. Epub 2014 Dec 4. Mol Cells. 2015. PMID: 25475546 Free PMC article.
-
The Genetics of Axon Guidance and Axon Regeneration in Caenorhabditis elegans.Genetics. 2016 Nov;204(3):849-882. doi: 10.1534/genetics.115.186262. Genetics. 2016. PMID: 28114100 Free PMC article. Review.
-
Neural regeneration in Caenorhabditis elegans.Annu Rev Genet. 2012;46:499-513. doi: 10.1146/annurev-genet-110711-155550. Epub 2012 Sep 4. Annu Rev Genet. 2012. PMID: 22974301 Free PMC article. Review.
Cited by
-
pxn-1 and pxn-2 May Interact Negatively during Neuronal Development and Aging in C. elegans.Mol Cells. 2015 Aug;38(8):729-33. doi: 10.14348/molcells.2015.0124. Epub 2015 Jul 21. Mol Cells. 2015. PMID: 26194821 Free PMC article.
-
Genetic Suppression of Basement Membrane Defects in Caenorhabditis elegans by Gain of Function in Extracellular Matrix and Cell-Matrix Attachment Genes.Genetics. 2018 Apr;208(4):1499-1512. doi: 10.1534/genetics.118.300731. Epub 2018 Feb 12. Genetics. 2018. PMID: 29440357 Free PMC article.
-
Bromide supplementation exacerbated the renal dysfunction, injury and fibrosis in a mouse model of Alport syndrome.PLoS One. 2017 Sep 5;12(9):e0183959. doi: 10.1371/journal.pone.0183959. eCollection 2017. PLoS One. 2017. PMID: 28873450 Free PMC article.
-
IGLR-2, a Leucine-Rich Repeat Domain Containing Protein, Is Required for the Host Defense in Caenorhabditis elegans.Front Immunol. 2020 Nov 30;11:561337. doi: 10.3389/fimmu.2020.561337. eCollection 2020. Front Immunol. 2020. PMID: 33329523 Free PMC article.
-
Molecular evolution, structure, and function of peroxidasins.Chem Biodivers. 2012 Sep;9(9):1776-93. doi: 10.1002/cbdv.201100438. Chem Biodivers. 2012. PMID: 22976969 Free PMC article.
References
-
- Alfonso T. B., Jones B. W. (2002). gcm2 promotes glial cell differentiation and is required with glial cells missing for macrophage development in Drosophila. Dev. Biol. 248, 369-383 - PubMed
-
- Andersen S. O. (1964). The cross-links in resilin identified as dityrosine and trityrosine. Biochim. Biophys. Acta 93, 213-215 - PubMed
-
- Benard C. Y., Boyanov A., Hall D. H., Hobert O. (2006). DIG-1, a novel giant protein, non-autonomously mediates maintenance of nervous system architecture. Development 133, 3329-3340 - PubMed
-
- Busch S. A., Silver J. (2007). The role of extracellular matrix in CNS regeneration. Curr. Opin. Neurobiol. 17, 120-127 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
