

Atoh1a expression must be restricted by Notch signaling for effective morphogenesis of the posterior lateral line primordium in zebrafish
- PMID: 20876657
- PMCID: PMC2947485
- DOI: 10.1242/dev.052761
Atoh1a expression must be restricted by Notch signaling for effective morphogenesis of the posterior lateral line primordium in zebrafish
Abstract
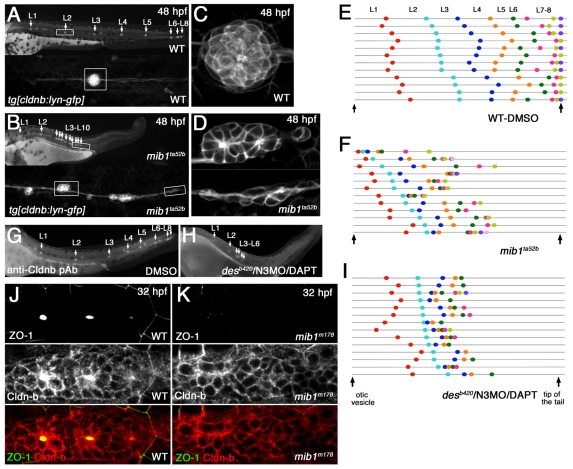
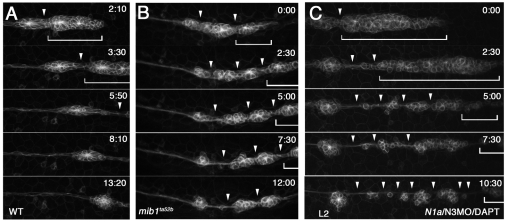
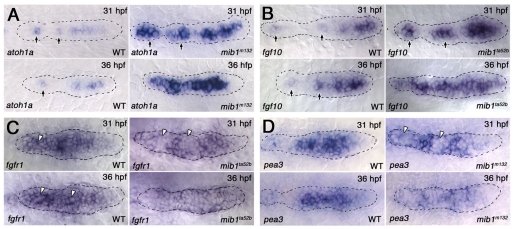
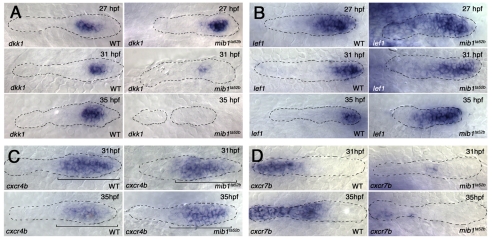
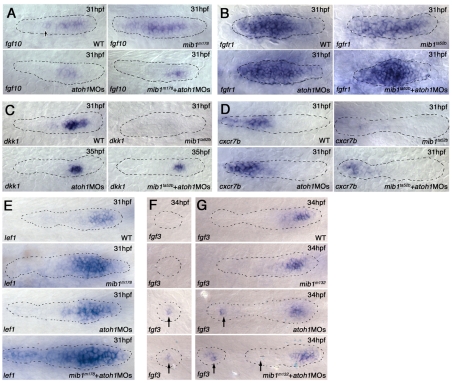
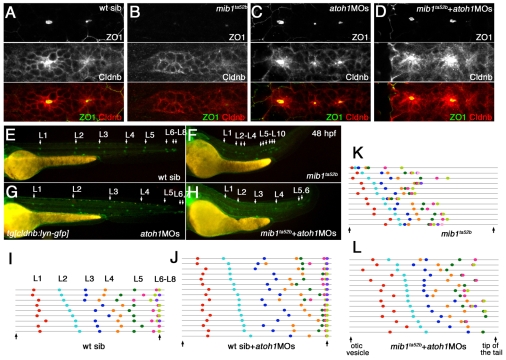
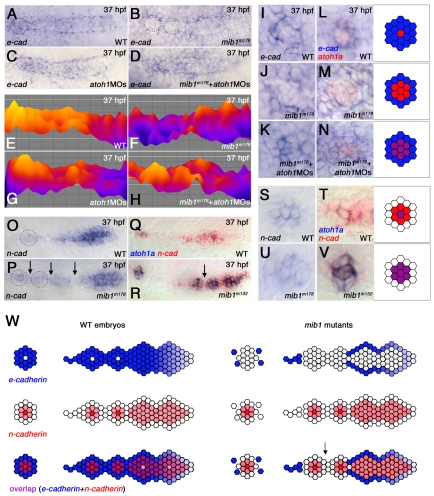
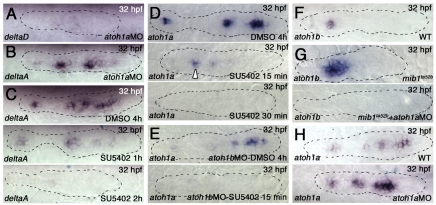
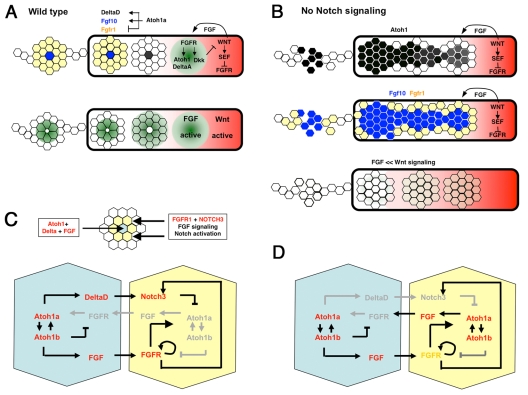
The posterior lateral line primordium (pLLp) migrates caudally, depositing neuromasts to establish the posterior lateral line system in zebrafish. A Wnt-dependent FGF signaling center at the leading end of the pLLp initiates the formation of `proneuromasts' by facilitating the reorganization of cells into epithelial rosettes and by initiating atoh1a expression. Expression of atoh1a gives proneuromast cells the potential to become sensory hair cells, and lateral inhibition mediated by Delta-Notch signaling restricts atoh1a expression to a central cell. We show that as atoh1a expression becomes established in the central cell, it drives expression of fgf10 and of the Notch ligand deltaD, while it inhibits expression of fgfr1. As a source of Fgf10, the central cell activates the FGF pathway in neighboring cells, ensuring that they form stable epithelial rosettes. At the same time, DeltaD activates Notch in neighboring cells, inhibiting atoh1a expression and ensuring that they are specified as supporting cells. When Notch signaling fails, unregulated atoh1a expression reduces Fgfr1 expression, eventually resulting in attenuated FGF signaling, which prevents effective maturation of epithelial rosettes in the pLLp. In addition, atoh1a inhibits e-cadherin expression, which is likely to reduce cohesion and contribute to fragmentation of the pLLp. Together, our observations reveal a genetic regulatory network that explains why atoh1a expression must be restricted by Notch signaling for effective morphogenesis of the pLLp.
Figures
Similar articles
-
Lef1 regulates Dusp6 to influence neuromast formation and spacing in the zebrafish posterior lateral line primordium.Development. 2013 Jun;140(11):2387-97. doi: 10.1242/dev.091348. Epub 2013 May 1. Development. 2013. PMID: 23637337 Free PMC article.
-
FGF-dependent mechanosensory organ patterning in zebrafish.Science. 2008 Jun 27;320(5884):1774-7. doi: 10.1126/science.1156547. Science. 2008. PMID: 18583612
-
Fgfr-Ras-MAPK signaling is required for apical constriction via apical positioning of Rho-associated kinase during mechanosensory organ formation.Development. 2012 Sep;139(17):3130-5. doi: 10.1242/dev.082271. Epub 2012 Jul 25. Development. 2012. PMID: 22833124 Free PMC article.
-
Signaling pathways regulating zebrafish lateral line development.Curr Biol. 2009 May 12;19(9):R381-6. doi: 10.1016/j.cub.2009.03.057. Curr Biol. 2009. PMID: 19439264 Review.
-
NetLogo agent-based models as tools for understanding the self-organization of cell fate, morphogenesis and collective migration of the zebrafish posterior Lateral Line primordium.Semin Cell Dev Biol. 2020 Apr;100:186-198. doi: 10.1016/j.semcdb.2019.12.015. Epub 2019 Dec 31. Semin Cell Dev Biol. 2020. PMID: 31901312 Review.
Cited by
-
Mib1 contributes to persistent directional cell migration by regulating the Ctnnd1-Rac1 pathway.Proc Natl Acad Sci U S A. 2017 Oct 31;114(44):E9280-E9289. doi: 10.1073/pnas.1712560114. Epub 2017 Oct 16. Proc Natl Acad Sci U S A. 2017. PMID: 29078376 Free PMC article.
-
Loss of ascl1a prevents secretory cell differentiation within the zebrafish intestinal epithelium resulting in a loss of distal intestinal motility.Dev Biol. 2013 Apr 15;376(2):171-86. doi: 10.1016/j.ydbio.2013.01.013. Epub 2013 Jan 23. Dev Biol. 2013. PMID: 23353550 Free PMC article.
-
The interplay of cell-cell and cell-substrate adhesion in collective cell migration.J R Soc Interface. 2014 Nov 6;11(100):20140684. doi: 10.1098/rsif.2014.0684. J R Soc Interface. 2014. PMID: 25165597 Free PMC article.
-
Notch signalling patterns retinal composition by regulating atoh7 during post-embryonic growth.Development. 2018 Nov 9;145(21):dev169698. doi: 10.1242/dev.169698. Development. 2018. PMID: 30337377 Free PMC article.
-
Cadherin-Mediated Cell Coupling Coordinates Chemokine Sensing across Collectively Migrating Cells.Curr Biol. 2019 Aug 5;29(15):2570-2579.e7. doi: 10.1016/j.cub.2019.06.061. Curr Biol. 2019. PMID: 31386838 Free PMC article.
References
-
- Abramoff M. D., Magelhaes P. J., Ram S. J. (2004). Image processing with ImageJ. Biophotonics Int. 11, 36-42
-
- Aman A., Piotrowski T. (2008). Wnt/beta-catenin and Fgf signaling control collective cell migration by restricting chemokine receptor expression. Dev. Cell 15, 749-761 - PubMed
-
- Cavallaro U., Christofori G. (2004). Cell adhesion and signalling by cadherins and Ig-CAMs in cancer. Nat. Rev. Cancer 4, 118-132 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
