

On the molecular basis of uracil recognition in DNA: comparative study of T-A versus U-A structure, dynamics and open base pair kinetics
- PMID: 20876689
- PMCID: PMC3025553
- DOI: 10.1093/nar/gkq812
On the molecular basis of uracil recognition in DNA: comparative study of T-A versus U-A structure, dynamics and open base pair kinetics
Abstract
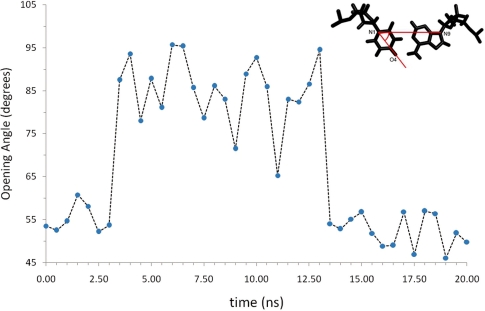
Uracil (U) can be found in DNA as a mismatch paired either to adenine (A) or to guanine (G). Removal of U from DNA is performed by a class of enzymes known as uracil-DNA-glycosylases (UDG). Recent studies suggest that recognition of U-A and U-G mismatches by UDG takes place via an extra-helical mechanism. In this work, we use molecular dynamics simulations to analyze the structure, dynamics and open base pair kinetics of U-A base pairs relative to their natural T-A counterpart in 12 dodecamers. Our results show that the presence of U does not alter the local conformation of B-DNA. Breathing dynamics and base pair closing kinetics are only weakly dependent on the presence of U versus T, with open T-A and U-A pairs lifetimes in the nanosecond timescale. Additionally, we observed spontaneous base flipping in U-A pairs. We analyze the structure and dynamics for this event and compare the results to available crystallographic data of open base pair conformations. Our results are in agreement with both structural and kinetic data derived from NMR imino proton exchange measurements, providing the first detailed description at the molecular level of elusive events such as spontaneous base pair opening and flipping in mismatched U-A sequences in DNA. Based on these results, we propose that base pair flipping can occur spontaneously at room temperature via a 3-step mechanism with an open base pair intermediate. Implications for the molecular basis of U recognition by UDG are discussed.
Figures








References
-
- Sousa MML, Krokan HE, Sluppaugh G. DNA-uracil and human pathology. Mol. Asp. Med. 2007;28:276–306. - PubMed
-
- Krokan HE, Drabløs F, Sluppaugh G. Uracil in DNA — occurrence, consequences and repair. Oncogene. 2002;21:8935–8948. - PubMed
-
- Friedberg EC, Walker GC, Siede W, Wood RD, Schultz RA, Ellenberger T. DNA Repair and Mutagenesis. Washington DC: ASM Press; 2006.
-
- Andersen S, Heine T, Sneve R, Konig I, Krokan HE, Epe B, Nilsen H. Incorporation of dUMP into DNA is a major source of spontaneous DNA damage, while excision of uracil is not required for cytotoxicity of fluoropyrimidines in mouse embryonic fibroblasts. Carcinogenesis. 2005;26:547–555. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
