

Neurofibromatosis-1 regulates neuroglial progenitor proliferation and glial differentiation in a brain region-specific manner
- PMID: 20876733
- PMCID: PMC2956210
- DOI: 10.1101/gad.1957110
Neurofibromatosis-1 regulates neuroglial progenitor proliferation and glial differentiation in a brain region-specific manner
Abstract
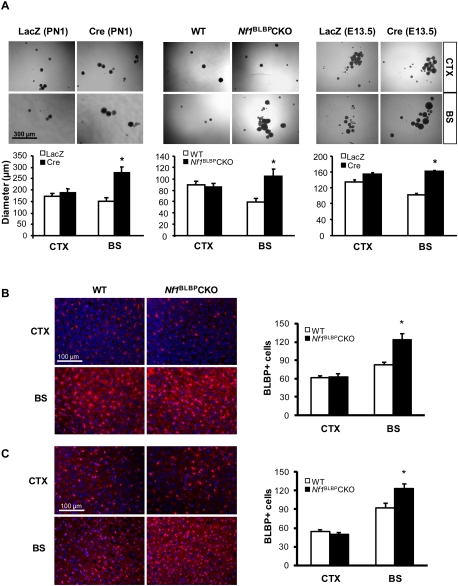
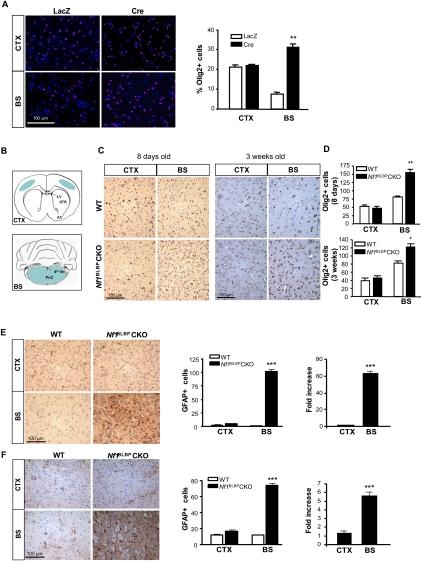
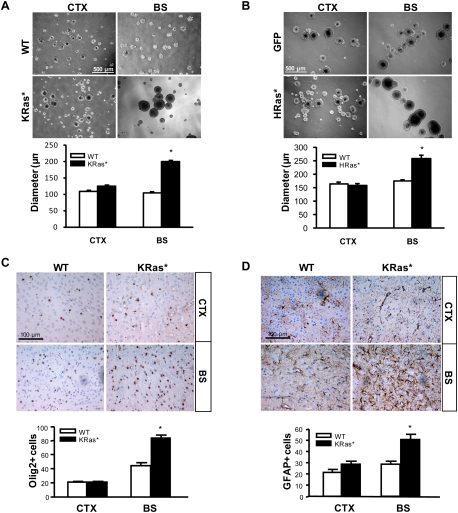
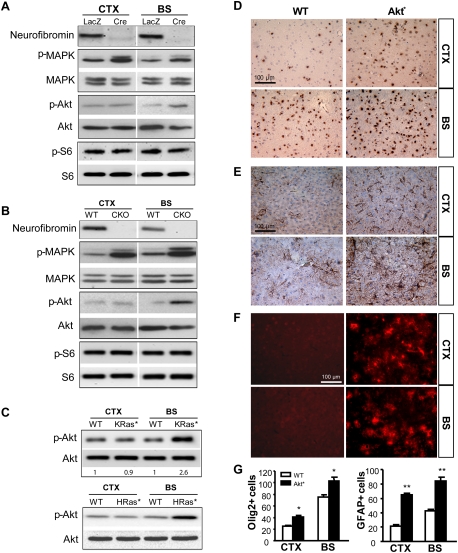
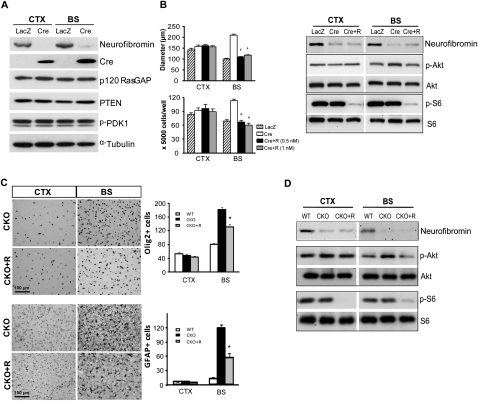
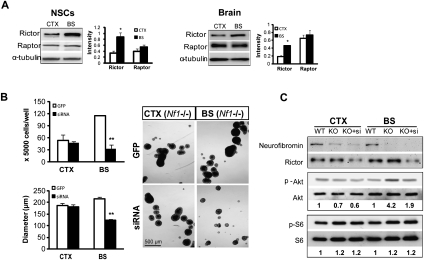
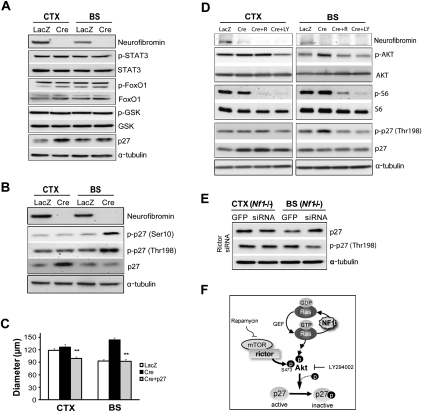
Recent studies have shown that neuroglial progenitor/stem cells (NSCs) from different brain regions exhibit varying capacities for self-renewal and differentiation. In this study, we used neurofibromatosis-1 (NF1) as a model system to elucidate a novel molecular mechanism underlying brain region-specific NSC functional heterogeneity. We demonstrate that Nf1 loss leads to increased NSC proliferation and gliogenesis in the brainstem, but not in the cortex. Using Nf1 genetically engineered mice and derivative NSC neurosphere cultures, we show that this brain region-specific increase in NSC proliferation and gliogenesis results from selective Akt hyperactivation. The molecular basis for the increased brainstem-specific Akt activation in brainstem NSCs is the consequence of differential rictor expression, leading to region-specific mammalian target of rapamycin (mTOR)/rictor-mediated Akt phosphorylation and Akt-regulated p27 phosphorylation. Collectively, these findings establish mTOR/rictor-mediated Akt activation as a key driver of NSC proliferation and gliogenesis, and identify a unique mechanism for conferring brain region-specific responses to cancer-causing genetic changes.
Figures
Comment in
-
Evidence for and against regional differences in neural stem and progenitor cells of the CNS.Genes Dev. 2010 Oct 15;24(20):2233-8. doi: 10.1101/gad.1988010. Genes Dev. 2010. PMID: 20952533 Free PMC article.
References
-
- Armando S, Lebrun A, Hugnot JP, Ripoll C, Saunier M, Simonneau L 2007. Neurosphere-derived neural cells show region-specific behaviour in vitro. Neuroreport 18: 1539–1542 - PubMed
-
- Basu TN, Gutmann DH, Fletcher JA, Glover TW, Collins FS, Downward J 1992. Aberrant regulation of ras proteins in malignant tumour cells from type 1 neurofibromatosis patients. Nature 356: 713–715 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous