

Cellular stress causes reversible, PRKAA1/2-, and proteasome-dependent ID2 protein loss in trophoblast stem cells
- PMID: 20876741
- PMCID: PMC3906848
- DOI: 10.1530/REP-10-0268
Cellular stress causes reversible, PRKAA1/2-, and proteasome-dependent ID2 protein loss in trophoblast stem cells
Abstract
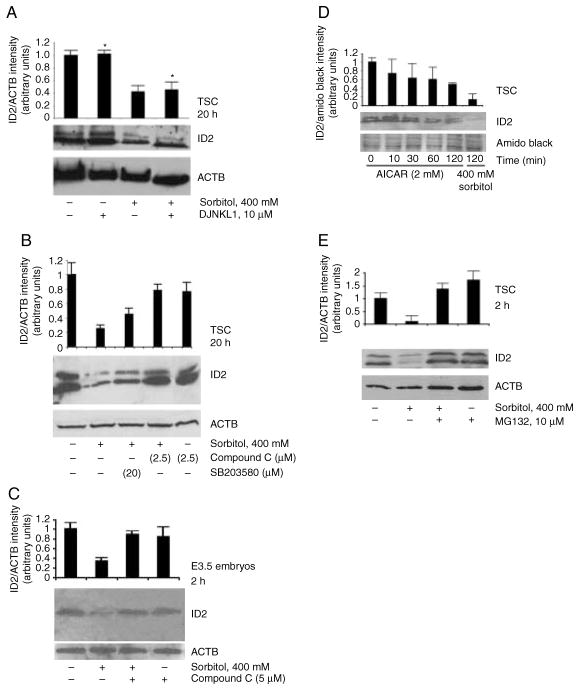
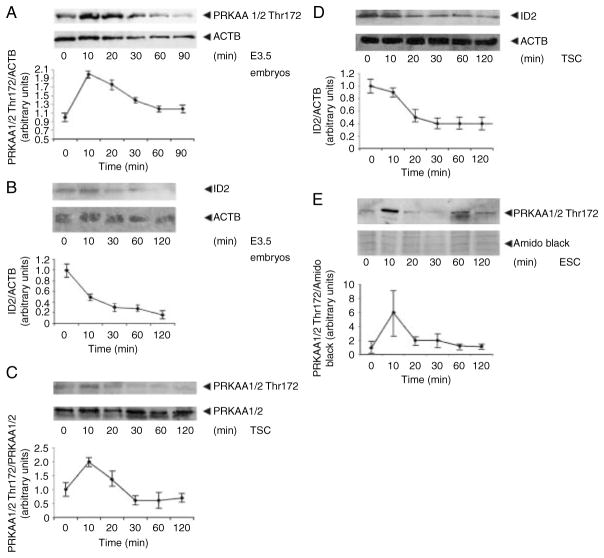
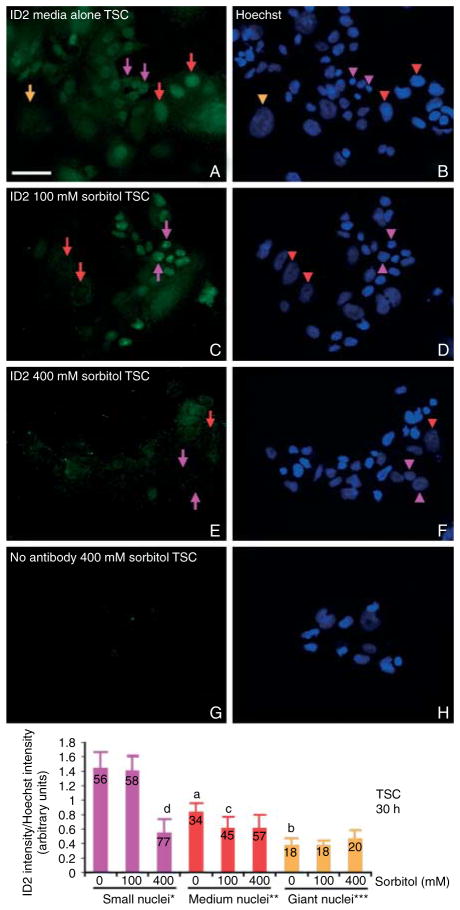
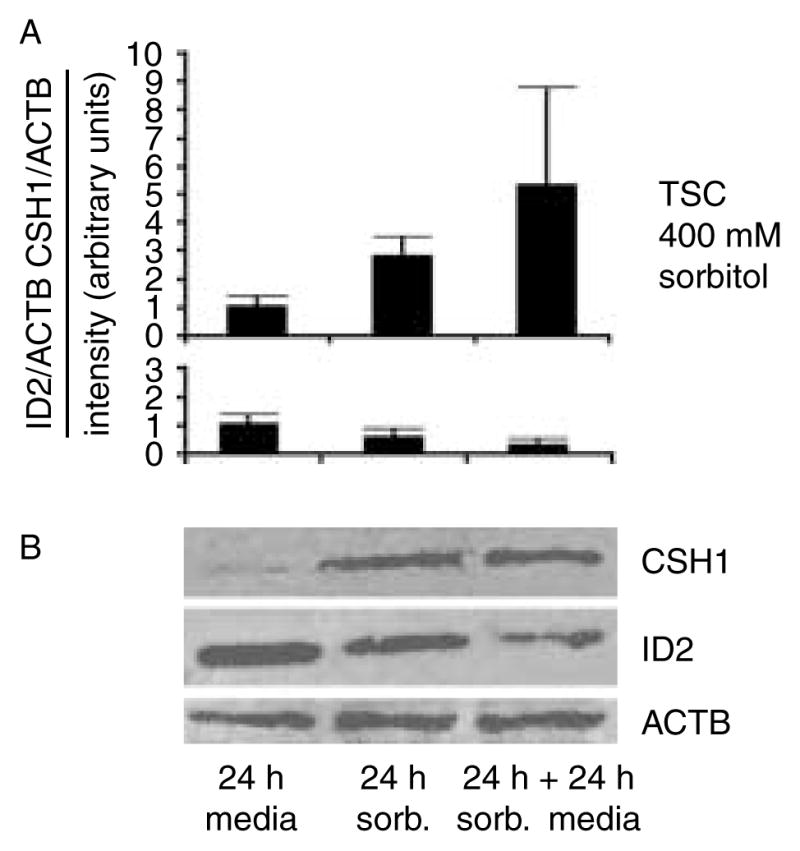
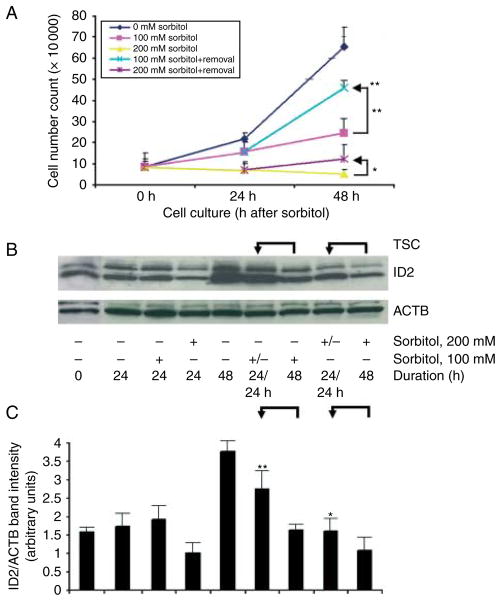
Stress reduces fertility, but the mechanisms mediating this are not understood. For a successful pregnancy, placental trophoblast stem cells (TSCs) in the implanting embryo proliferate and then a subpopulation differentiates to produce hormones. Normally, differentiation occurs when inhibitor of differentiation 2 (ID2) protein is lost in human and mouse placental stem cells. We hypothesize that stress enzyme-dependent differentiation occurs in association with insufficient TSC accumulation. We studied a well-defined model where TSC differentiation requires ID2 loss. The loss of ID2 derepresses the promoter of chorionic somatomammotropin hormone 1 (CSH1), the first hormone after implantation. Csh1 mRNA is known to be induced in stressed TSCs. In this study, we demonstrate that AMP-activated protein kinase (PRKAA1/2, aka AMPK) mediates the stress-induced proteasome-dependent loss of ID2 at high stress levels. At very low stress levels, PRKAA1/2 mediates metabolic adaptation exemplified by the inactivation of acetyl coA carboxylase by phosphorylation without ID2 loss. At the highest stress levels, irreversible TSC differentiation as defined by ID2 loss and slower cell accumulation occurs. However, lower stress levels lead to reversible differentiation accompanied by metabolic adaptation. These data support the hypothesis that PRKAA1/2 mediates preparation for differentiation that is induced by stress at levels where a significant decrease in cell accumulation occurs. This supports the interpretation that enzyme-mediated increases in differentiation may compensate when insufficient numbers of stem cells accumulate.
Conflict of interest statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.
Figures

References
-
- Bell CE, Lariviere NM, Watson PH, Watson AJ. Mitogen-activated protein kinase (MAPK) pathways mediate embryonic responses to culture medium osmolarity by regulating Aquaporin 3 and 9 expression and localization, as well as embryonic apoptosis. Human Reproduction. 2009;24:1373–1386. 10.1093/humrep/dep010. - PubMed
-
- Chai N, Patel Y, Jacobson K, McMahon J, McMahon A, Rappolee DA. FGF is an essential regulator of the fifth cell division in preimplantation mouse embryos. Developmental Biology. 1998;198:105–115. 10.1016/S0012-1606(98)80031-6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
