

Epigenetic regulation of repetitive elements is attenuated by prolonged heat stress in Arabidopsis
- PMID: 20876829
- PMCID: PMC2965555
- DOI: 10.1105/tpc.110.078493
Epigenetic regulation of repetitive elements is attenuated by prolonged heat stress in Arabidopsis
Abstract
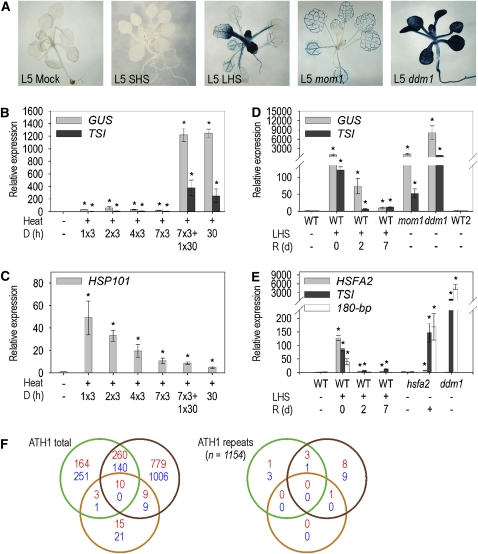
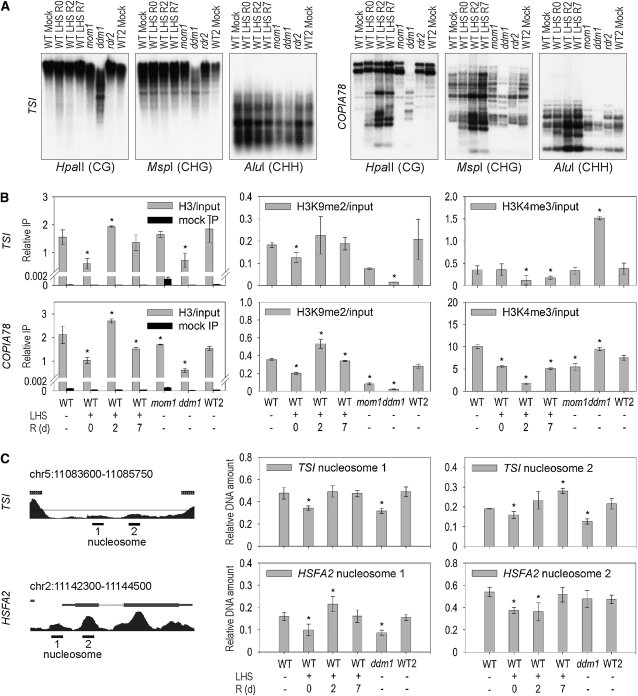
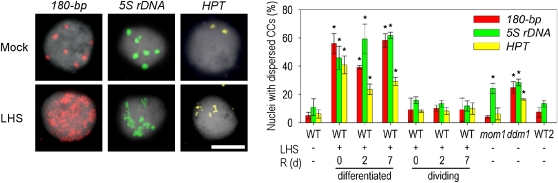
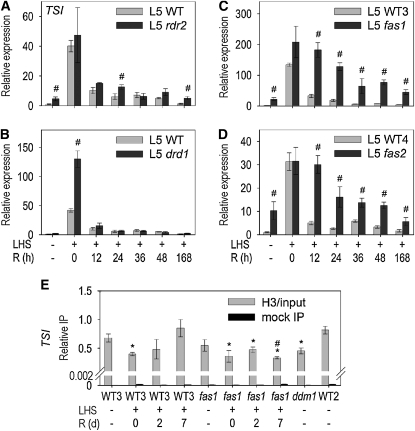
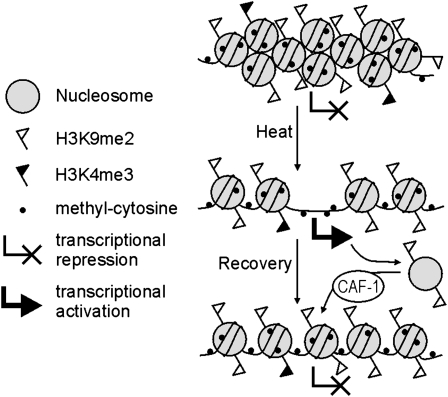
Epigenetic factors determine responses to internal and external stimuli in eukaryotic organisms. Whether and how environmental conditions feed back to the epigenetic landscape is more a matter of suggestion than of substantiation. Plants are suitable organisms with which to address this question due to their sessile lifestyle and diversification of epigenetic regulators. We show that several repetitive elements of Arabidopsis thaliana that are under epigenetic regulation by transcriptional gene silencing at ambient temperatures and upon short term heat exposure become activated by prolonged heat stress. Activation can occur without loss of DNA methylation and with only minor changes to histone modifications but is accompanied by loss of nucleosomes and by heterochromatin decondensation. Whereas decondensation persists, nucleosome loading and transcriptional silencing are restored upon recovery from heat stress but are delayed in mutants with impaired chromatin assembly functions. The results provide evidence that environmental conditions can override epigenetic regulation, at least transiently, which might open a window for more permanent epigenetic changes.
Figures
References
-
- Amedeo P., Habu Y., Afsar K., Mittelsten Scheid O., Paszkowski J. (2000). Disruption of the plant gene MOM releases transcriptional silencing of methylated genes. Nature 405: 203–206 - PubMed
-
- Bilang R., Iida S., Peterhans A., Potrykus I., Paszkowski J. (1991). The 3′-terminal region of the hygromycin-B-resistance gene is important for its activity in Escherichia coli and Nicotiana tabacum. Gene 100: 247–250 - PubMed
-
- Binelli G., Mascarenhas J. (1990). Arabidopsis: Sensitivity of growth to high temperature. Dev. Genet. 11: 294–298
-
- Chan S.W., Henderson I.R., Jacobsen S.E. (2005). Gardening the genome: DNA methylation in Arabidopsis thaliana. Nat. Rev. Genet. 6: 351–360 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
