

A β(IV)-spectrin/CaMKII signaling complex is essential for membrane excitability in mice
- PMID: 20877009
- PMCID: PMC2947241
- DOI: 10.1172/JCI43621
A β(IV)-spectrin/CaMKII signaling complex is essential for membrane excitability in mice
Abstract
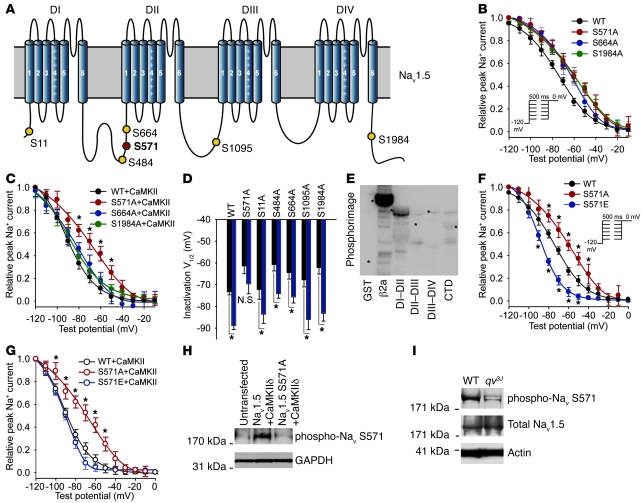
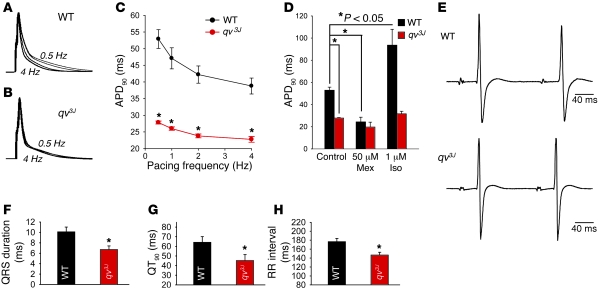
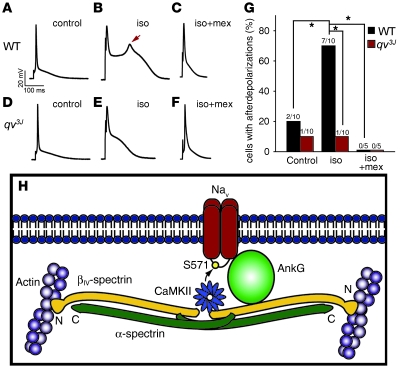
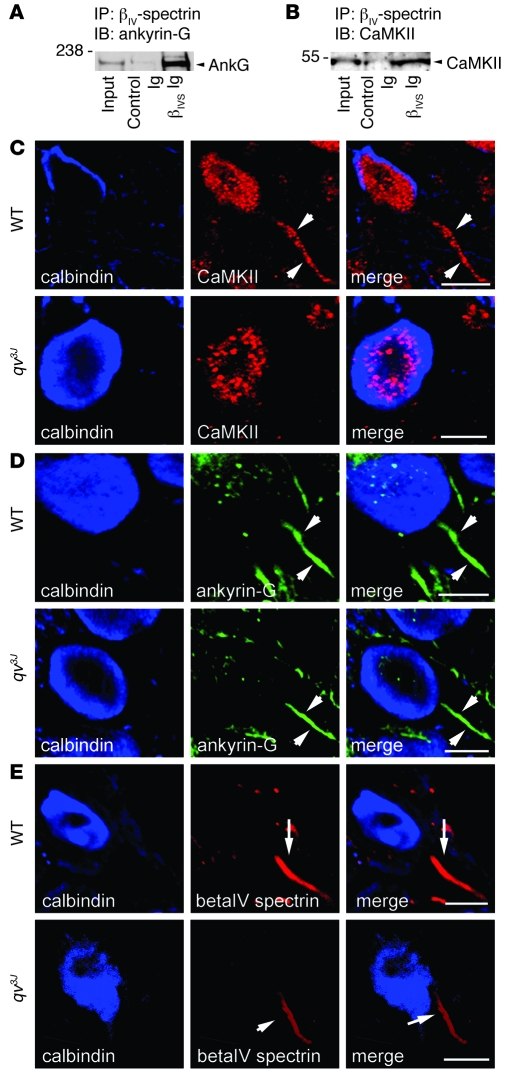
Ion channel function is fundamental to the existence of life. In metazoans, the coordinate activities of voltage-gated Na(+) channels underlie cellular excitability and control neuronal communication, cardiac excitation-contraction coupling, and skeletal muscle function. However, despite decades of research and linkage of Na(+) channel dysfunction with arrhythmia, epilepsy, and myotonia, little progress has been made toward understanding the fundamental processes that regulate this family of proteins. Here, we have identified β(IV)-spectrin as a multifunctional regulatory platform for Na(+) channels in mice. We found that β(IV)-spectrin targeted critical structural and regulatory proteins to excitable membranes in the heart and brain. Animal models harboring mutant β(IV)-spectrin alleles displayed aberrant cellular excitability and whole animal physiology. Moreover, we identified a regulatory mechanism for Na(+) channels, via direct phosphorylation by β(IV)-spectrin-targeted calcium/calmodulin-dependent kinase II (CaMKII). Collectively, our data define an unexpected but indispensable molecular platform that determines membrane excitability in the mouse heart and brain.
Figures
Comment in
-
Location, location, regulation: a novel role for β-spectrin in the heart.J Clin Invest. 2010 Oct;120(10):3434-7. doi: 10.1172/JCI44810. Epub 2010 Sep 27. J Clin Invest. 2010. PMID: 20877007 Free PMC article.
Similar articles
-
Location, location, regulation: a novel role for β-spectrin in the heart.J Clin Invest. 2010 Oct;120(10):3434-7. doi: 10.1172/JCI44810. Epub 2010 Sep 27. J Clin Invest. 2010. PMID: 20877007 Free PMC article.
-
Ankyrin-G coordinates intercalated disc signaling platform to regulate cardiac excitability in vivo.Circ Res. 2014 Nov 7;115(11):929-38. doi: 10.1161/CIRCRESAHA.115.305154. Epub 2014 Sep 19. Circ Res. 2014. PMID: 25239140 Free PMC article.
-
Ca2+/calmodulin-dependent protein kinase II (CaMKII) regulates cardiac sodium channel NaV1.5 gating by multiple phosphorylation sites.J Biol Chem. 2012 Jun 8;287(24):19856-69. doi: 10.1074/jbc.M111.322537. Epub 2012 Apr 18. J Biol Chem. 2012. PMID: 22514276 Free PMC article.
-
Ca2+-dependent modulation of voltage-gated myocyte sodium channels.Biochem Soc Trans. 2021 Nov 1;49(5):1941-1961. doi: 10.1042/BST20200604. Biochem Soc Trans. 2021. PMID: 34643236 Free PMC article. Review.
-
Calcium/calmodulin-dependent kinase II regulation of cardiac ion channels.J Cardiovasc Pharmacol. 2009 Sep;54(3):180-7. doi: 10.1097/FJC.0b013e3181a25078. J Cardiovasc Pharmacol. 2009. PMID: 19333131 Free PMC article. Review.
Cited by
-
New therapeutic targets in cardiology: arrhythmias and Ca2+/calmodulin-dependent kinase II (CaMKII).Circulation. 2012 Oct 23;126(17):2125-39. doi: 10.1161/CIRCULATIONAHA.112.124990. Circulation. 2012. PMID: 23091085 Free PMC article. Review. No abstract available.
-
Defects in cytoskeletal signaling pathways, arrhythmia, and sudden cardiac death.Front Physiol. 2012 May 3;3:122. doi: 10.3389/fphys.2012.00122. eCollection 2012. Front Physiol. 2012. PMID: 22586405 Free PMC article.
-
The promise of CaMKII inhibition for heart disease: preventing heart failure and arrhythmias.Expert Opin Ther Targets. 2013 Aug;17(8):889-903. doi: 10.1517/14728222.2013.809064. Epub 2013 Jun 24. Expert Opin Ther Targets. 2013. PMID: 23789646 Free PMC article. Review.
-
CaMKII in Regulation of Cell Death During Myocardial Reperfusion Injury.Front Mol Biosci. 2021 Jun 1;8:668129. doi: 10.3389/fmolb.2021.668129. eCollection 2021. Front Mol Biosci. 2021. PMID: 34141722 Free PMC article. Review.
-
Spectrin-based skeleton as an actor in cell signaling.Cell Mol Life Sci. 2012 Jan;69(2):191-201. doi: 10.1007/s00018-011-0804-5. Epub 2011 Aug 30. Cell Mol Life Sci. 2012. PMID: 21877118 Free PMC article. Review.
References
-
- Zhang R, et al. Calmodulin kinase II inhibition protects against structural heart disease. Nat Med. 2005;11(4):409–417. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL079031/HL/NHLBI NIH HHS/United States
- K99 HL096805/HL/NHLBI NIH HHS/United States
- HL70250/HL/NHLBI NIH HHS/United States
- R01 HL096652/HL/NHLBI NIH HHS/United States
- R01 HL084583/HL/NHLBI NIH HHS/United States
- HL096805/HL/NHLBI NIH HHS/United States
- R01 HL070250/HL/NHLBI NIH HHS/United States
- R00 HL096805/HL/NHLBI NIH HHS/United States
- NS044916/NS/NINDS NIH HHS/United States
- UL1 RR024979/RR/NCRR NIH HHS/United States
- R37 NS044916/NS/NINDS NIH HHS/United States
- HL084583/HL/NHLBI NIH HHS/United States
- R01 NS044916/NS/NINDS NIH HHS/United States
- HL62494/HL/NHLBI NIH HHS/United States
- R01 HL062494/HL/NHLBI NIH HHS/United States
- R01 HL083422/HL/NHLBI NIH HHS/United States
- HL083422/HL/NHLBI NIH HHS/United States
- HL079031/HL/NHLBI NIH HHS/United States
- T32 HL007121/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
