

doi: 10.1039/BK9781847559159-00053.
Epub 2009 Jan 30.
Cobalamin- and corrinoid-dependent enzymes
Affiliations
- PMID: 20877792
- PMCID: PMC3120101
- DOI: 10.1039/BK9781847559159-00053
Item in Clipboard
Cobalamin- and corrinoid-dependent enzymes
Met Ions Life Sci.
2009.
Abstract
This chapter reviews the literature on cobalamin- and corrinoid-containing enzymes. These enzymes fall into two broad classes, those using methylcobalamin or related methylcorrinoids as prosthetic groups and catalyzing methyl transfer reactions, and those using adenosylcobalamin as the prosthetic group and catalyzing the generation of substrate radicals that in turn undergo rearrangements and/or eliminations.
Figures
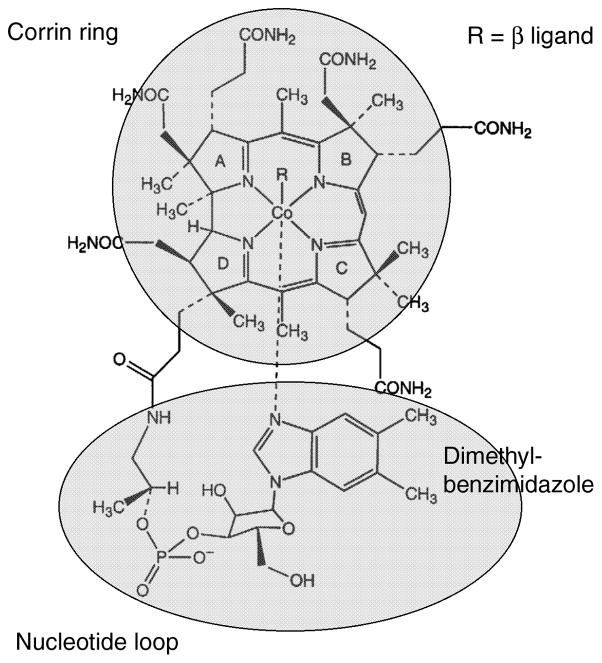
R is a methyl group in methylcobalamin or an adenosyl group in adenosylcobalamin.
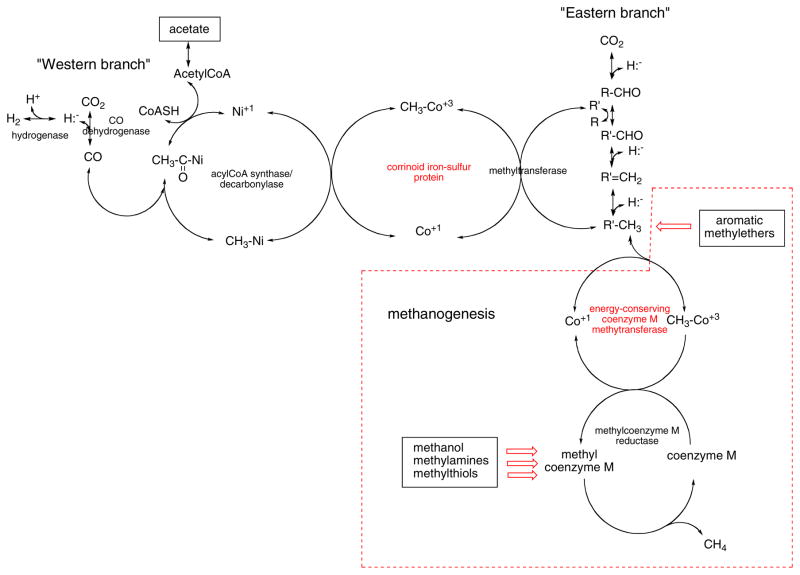
The corrinoid methyl transferases are labeled in red. Red arrows indicate pathways, not shown in detail, that also involve corrinoid proteins. The boxed reactions are not part of the Wood-Ljungdahl pathway but are unique to methanogens. R′ is tetrahydrofolate or a tetrahydropterin analogue of tetrahydrofolate. The charge on the nickel that serves as the methyl acceptor on acylCoA synthase/decarbonylase is shown as +1, but whether it is Ni+1 or Ni0 rremains a matter for debate , .
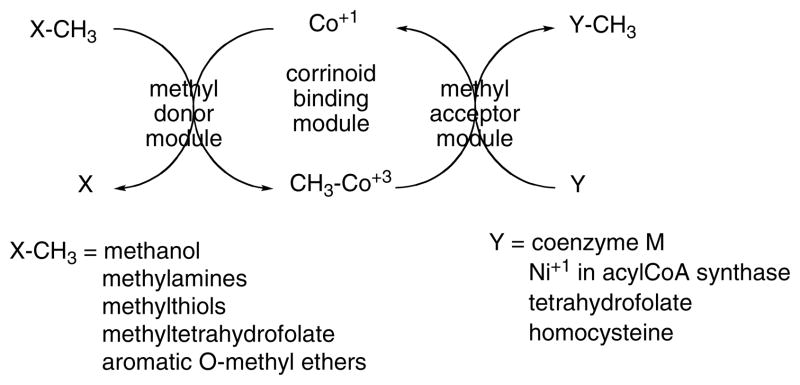
Basic pattern for corrinoid methyltransferases.
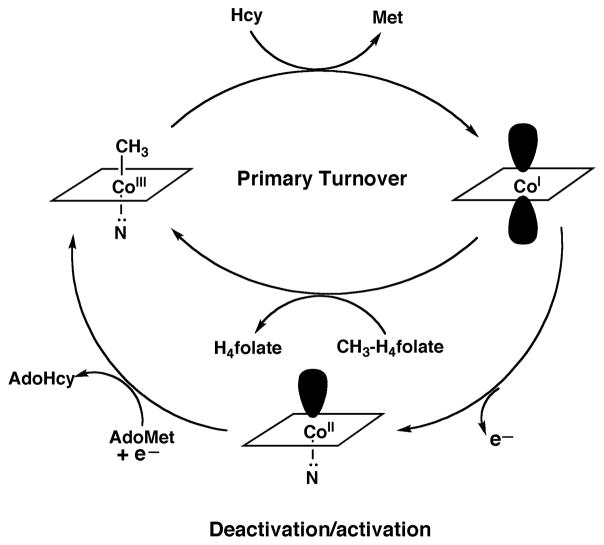
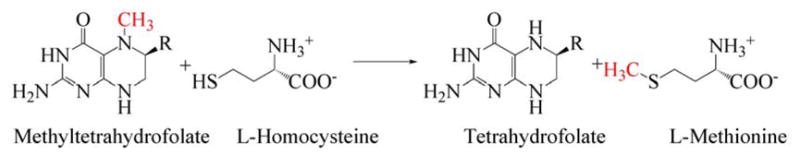
During primary turnover, the enzyme-bound cobalamin cycles between methylcobalamin and cob(I)alamin forms as the prosthetic group is alternately methylated by methyltetrahydrofolate (CH3-H4folate) and demethylated by transfer of the methyl group to homocysteine (Hcy). During turnover under microaerophilic conditions, the cob(I)alamin form of the enzyme is oxidized to cob(II)alamin about once in every 2000 turnovers. This form of the enzyme is inactive, and reactivation requires a reductive methylation in which the reduction of cob(II)alamin to cob(I)alamin is coupled to a highly exergonic methylation using adenosylmethionine (AdoMet) as the methyl donor.
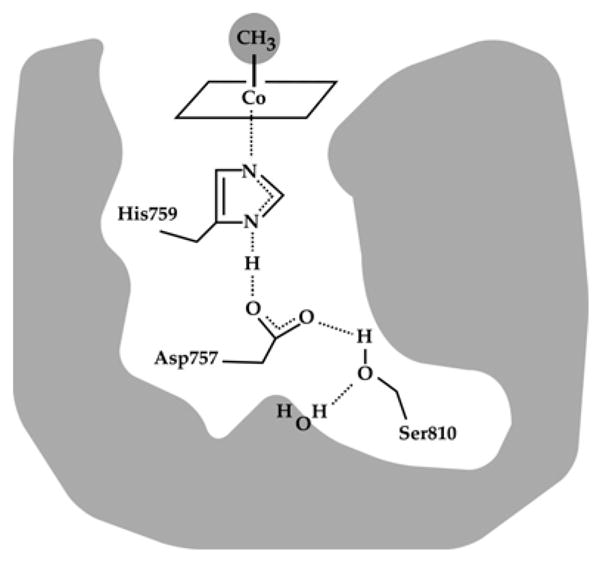
A hydrogen bonded network comprising His 759, Asp757 and Ser810 links His759 to the external solvent; these three residues which are absolutely conserved in all MetH enzymes, are referred to as the ligand triad. (Reprinted with permission, from )
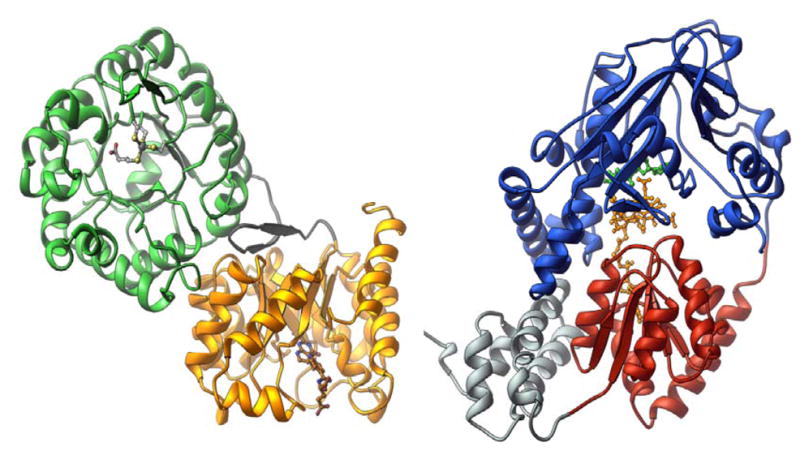
The structure of the homocysteine-binding (green) and folate-binding (gold) modules of MetH was determined with protein from Thermotoga maritima , and the structure of the cobalamin-binding (red) and reactivation (blue) modules of MetH was determined with the His759Gly mutant of the enzyme from Escherichia coli . The cobalamin-binding module also contains a four helix bundle, referred to as the “cap” and shown in grey.
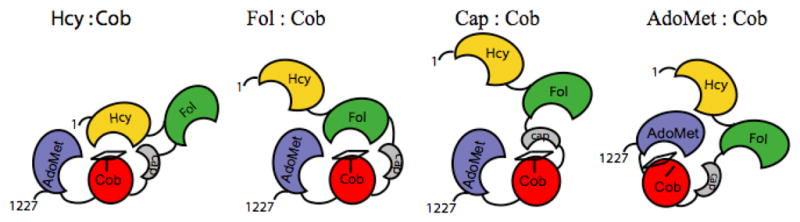
The four modules are shown in gold (Hcy-binding), green (folate-binding), red (cobalamin-binding domain) and gray (cap domain), and blue (AdoMet-binding). The corrin ring of methylcobalamin is indicated by the rectangle on top of the cobalamin-binding domain, and His759 is indicated by the vertical line. In the AdoMet:Cob conformation, the histidine is displaced as indicated in the cartoon, and the corrin ring tilts away from the cobalamin-binding domain and from Cα of His 759. Reprinted from .
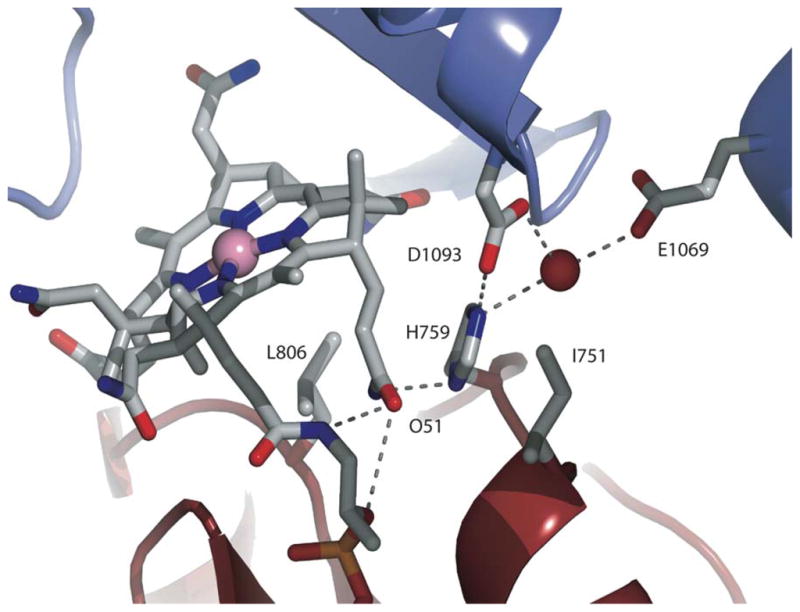
Nε2 of His759 interacts with the AdoMet module directly through a hydrogen bond to Asp1093 and via a water-mediated hydrogen bond to Glu1069. Nδ1 of His759 forms a hydrogen bond with the amide of the propionamide side chain of ring B of the cobalamin (not shown). Adapted from .
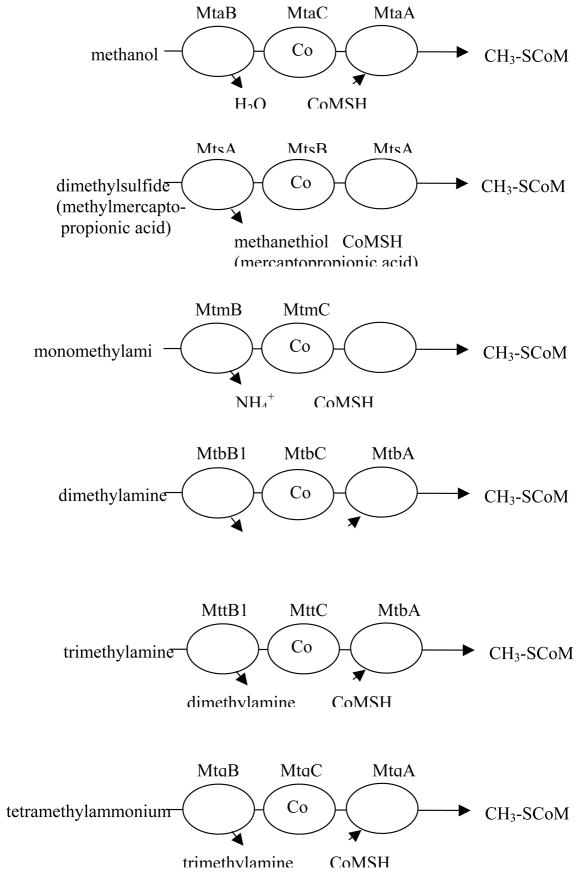
Complexes catalyzing methyl transfer from methyl donors to coenzyme M. The gene product designations are shown above each protein. The first two letters, mt, indicate involvement of the gene product in methyl transfer, the third letter indicates the substrate: a for methanol, s for methylthiols, m for monomethylamine, b for dimethylamine, t for trimethylamine and q for tetramethylammonium. The final letter designates the polypeptide function: where B is the substrate-specific methyltransferase that methylates the corrinoid protein with a methyl group derived from substrate, C is the corrinoid binding polypeptide and A is the CoM-methylating protein, as originally suggested by Krzycki and coworkers .
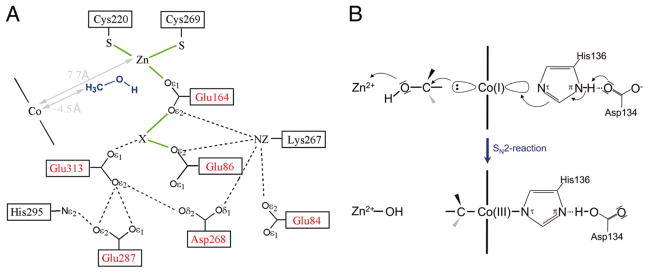
Reproduced, with permission, from . The electron density indicated by X has been modeled as a potassium ion.
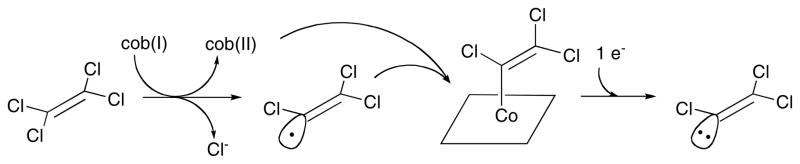
Redrawn, with permission from .
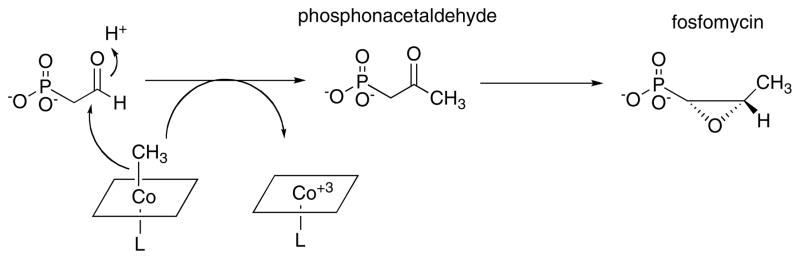
Adapted with permission from .
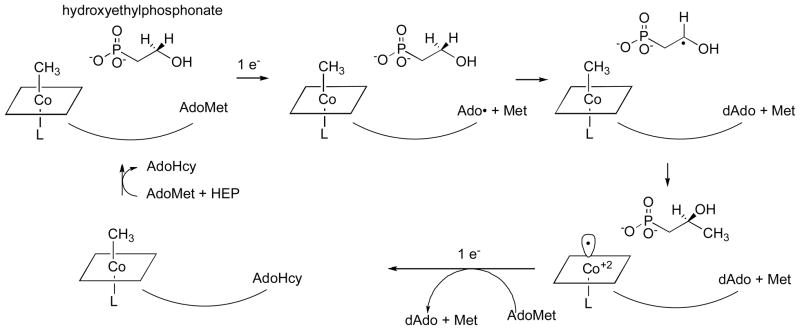
Adapted with permission from
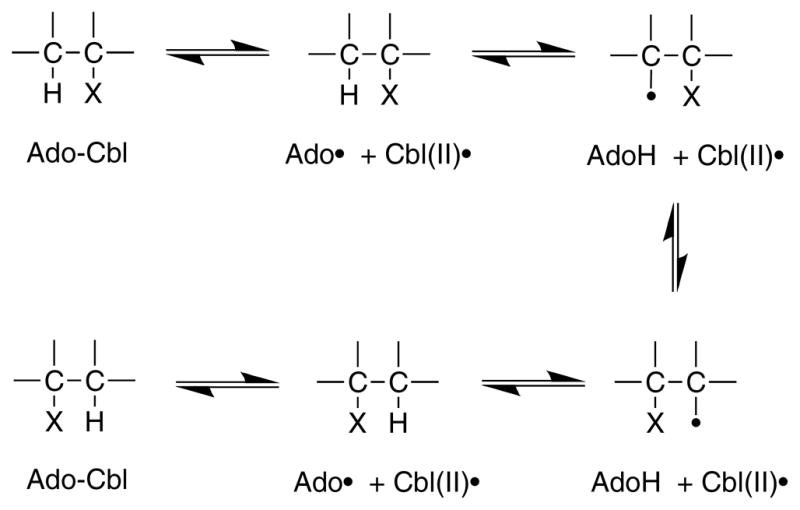
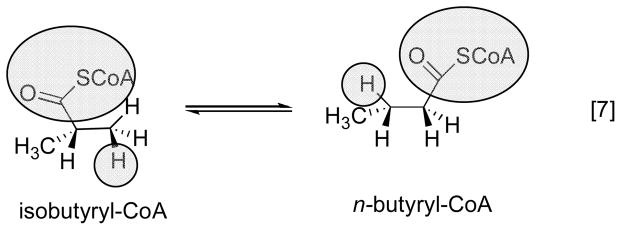
Basic mechanism of AdoCbl-dependent rearrangements
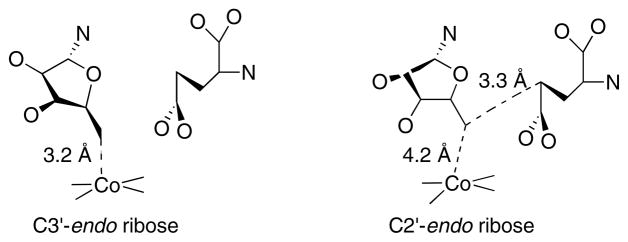
On the left the ribose assumes a C3′-endo conformation, placing the 5′-carbon within bonding distance of the cobalt of cobalamin and distant from Cγ of the glutamate substrate. On the right the ribose assumes a C2′-endo conformation, leading to breakage of the carbon-cobalt bond and placing C5′ within van der Waal’s distance of Cγ of the substrate.
The adenosyl radical formed by cleavage of the C-Co bond of AdoCbl abstracts a hydrogen from glutamate to yield the glutamyl radical. This leads to the elimination of acrylate and formation of glycyl radical, which in turn condenses with acrylate to form the β-methylaspartate radical. In the final step, the product radical abstracts hydrogen from 5′-deoxyadenosine to regenerate the adenosyl radical, which can now recombine with cob(II)alamin to reform AdoCbl.
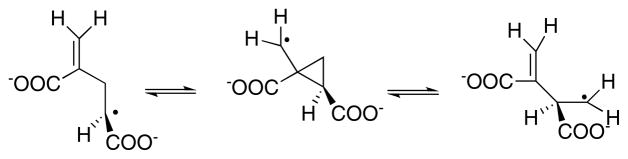
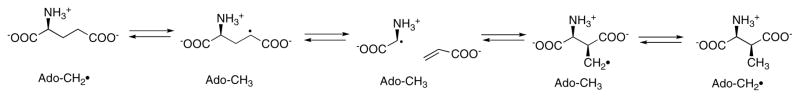
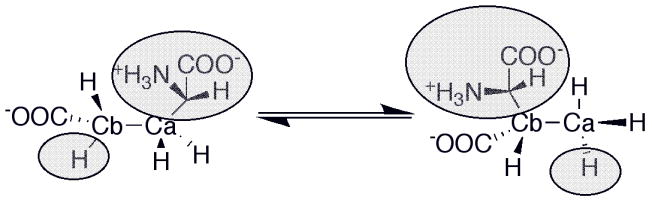
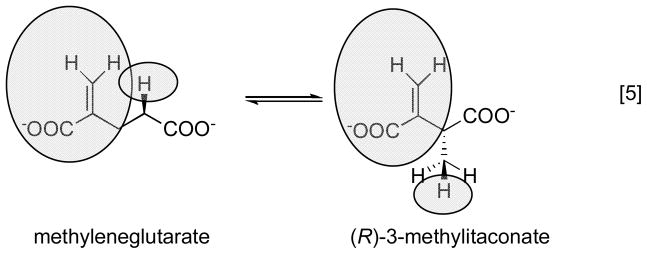
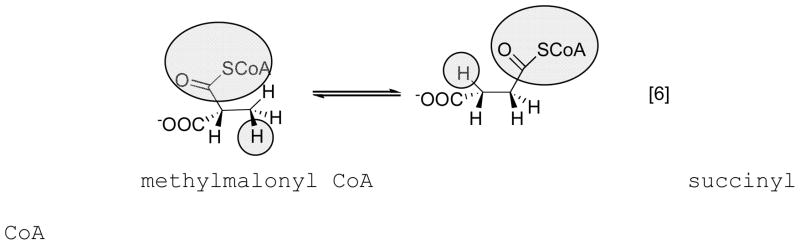
Because the migrating carbon is sp2, a cyclopropylcarbinyl radical can form to mediate the transfer of the radical between Cα and Cβ.
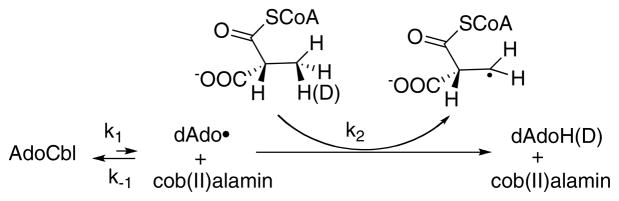
A rapid but unfavorable equilibrium between AdoCbl and the homolytically cleaved dAdo radical and cob(II)alamin precedes a rate-limiting hydrogen atom abstraction from the substrate .
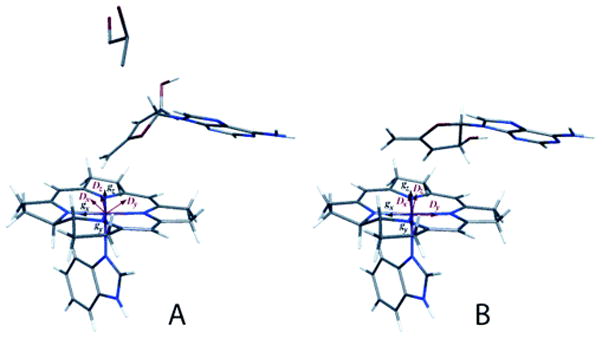
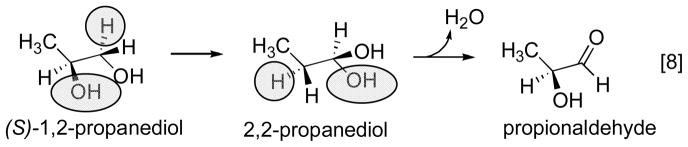
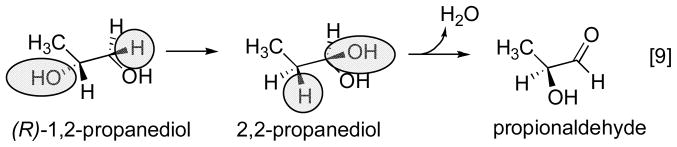
Reprinted, with permission from . Electron paramagnetic analysis of diol dehydrase reconstituted with 3′,4′-anhydroAdoCbl and subjected to homolytic cleavage to form the anhydroadenosyl radical and cob(II)alamin in the presence (A) and absence (B) of (R,S)-1,2-propanediol was used to model the structures. In the absence of substrate, the anhydroribosyl moity rotates about 60° relative to its position in the presence of substrate, bringing the radical into a position that would be appropiate for hydrogen atom abstraction from substrate and that would not permit formation of a carbon-cobalt bond.
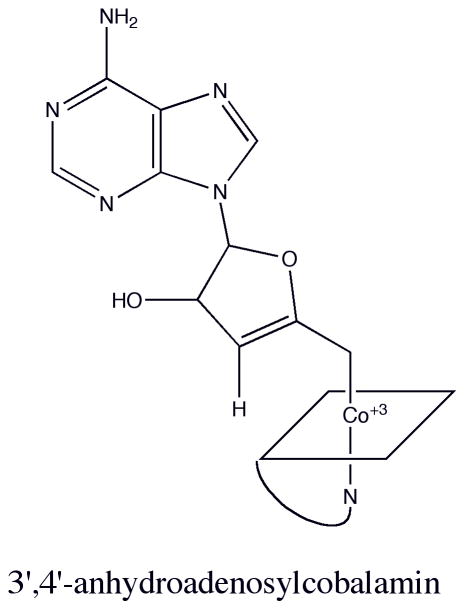
Structure of 3′,4′-anhydroadenosylcobalamin.
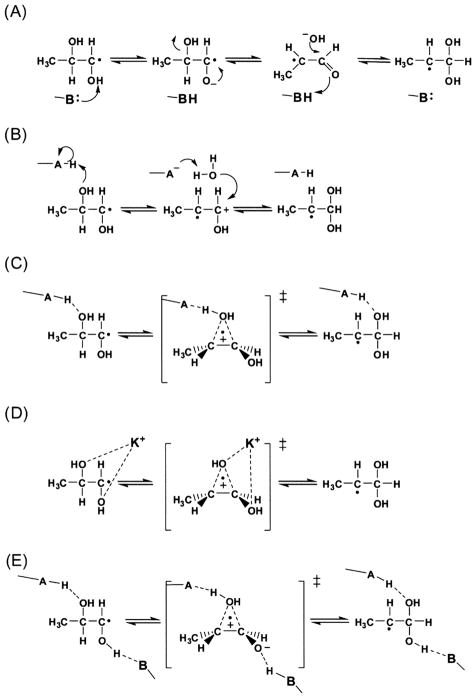
Reprinted with permission from Schwartz et al. . The mechanisms proposed involve (a) general base catalysis, (b) general acid catalysis, (c) partial protonation (hydrogen bonding) (d) electrophilic catalysis by the activating potassium ion, or (e) combined general acid/base (push-pull) catalysis.
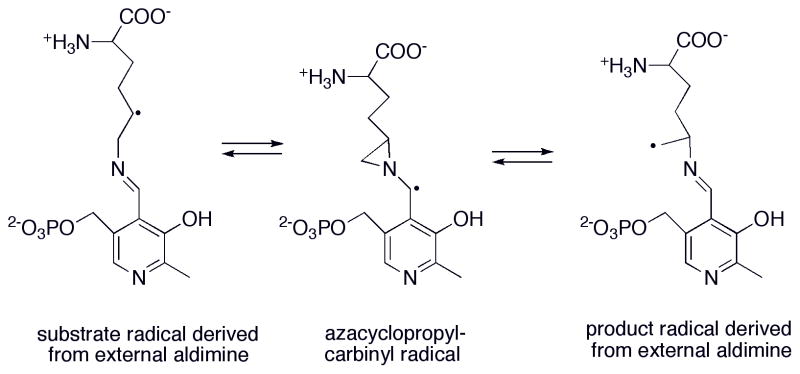
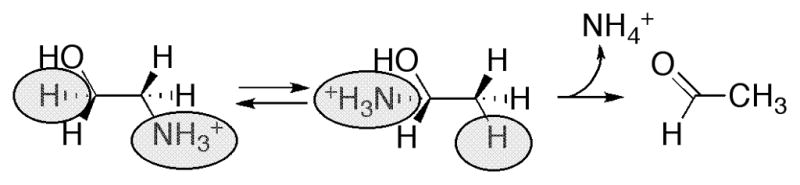
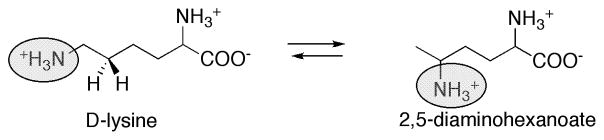
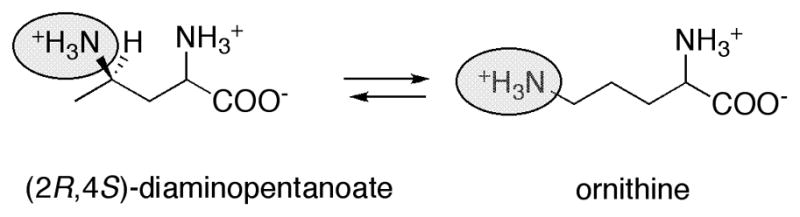
Proposed role for pyridoxal phosphate in the reaction catalyzed by lysine 5,6-aminomutase.
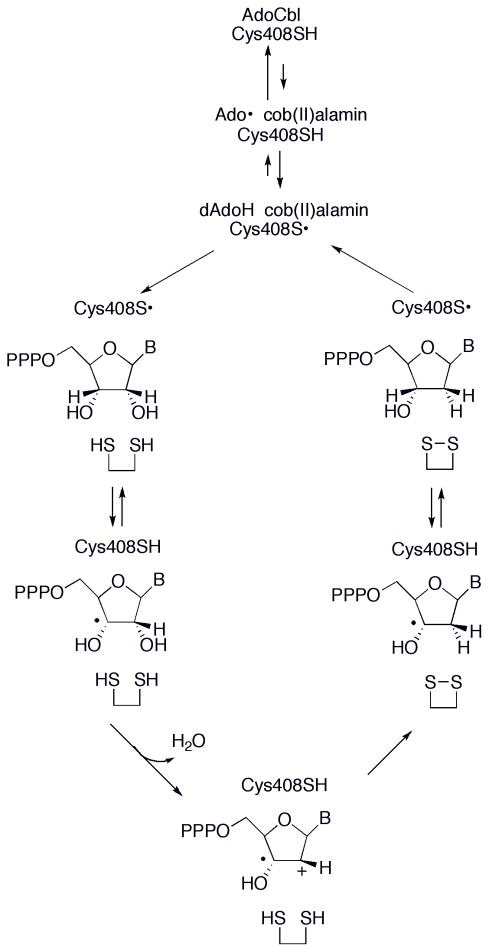
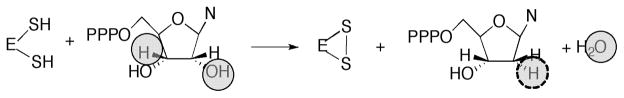
Proposed mechanism for AdoCbl-dependent ribonucleotide reductase.

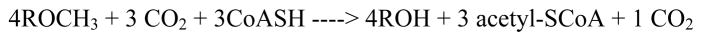
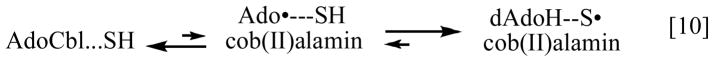

References
-
- Ragsdale SW, Lindahl PA, Munck E. J Biol Chem. 1987;262:14289–14297. - PubMed
-
- Stupperich E, Eisenger HJ, Albracht SPJ. Eur J Biochem. 1990;193:105–109. - PubMed
-
- Ragsdale SW. In: Chemistry and biochemistry of B12. Banerjee R, editor. John Wiley; New York: 1999. pp. 633–653.
-
- Banerjee RV, Johnston NL, Sobeski JK, Datta P, Matthews RG. J Biol Chem. 1989;264:13888–13895. - PubMed
-
- Amaratunga M, Fluhr K, Jarrett JT, Drennan CL, Ludwig ML, Matthews RG, Scholten JD. Biochemistry. 1996;35:2453–2463. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources