

The solution structure of a plant calmodulin and the CaM-binding domain of the vacuolar calcium-ATPase BCA1 reveals a new binding and activation mechanism
- PMID: 20880850
- PMCID: PMC2992282
- DOI: 10.1074/jbc.M110.131201
The solution structure of a plant calmodulin and the CaM-binding domain of the vacuolar calcium-ATPase BCA1 reveals a new binding and activation mechanism
Abstract
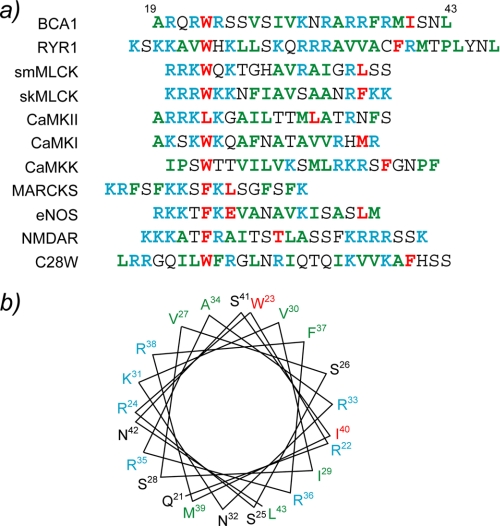
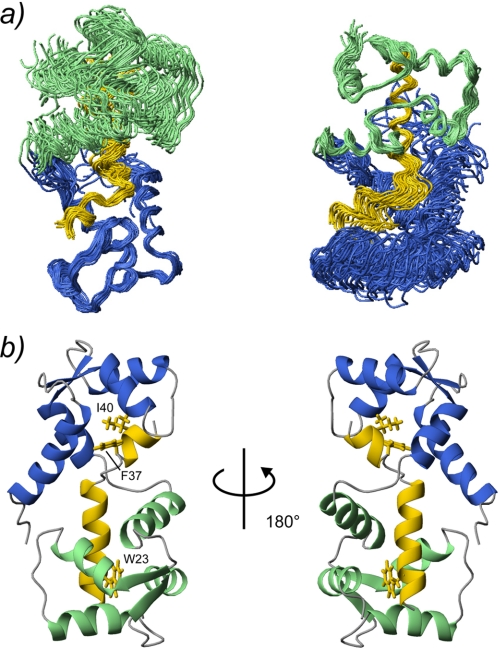
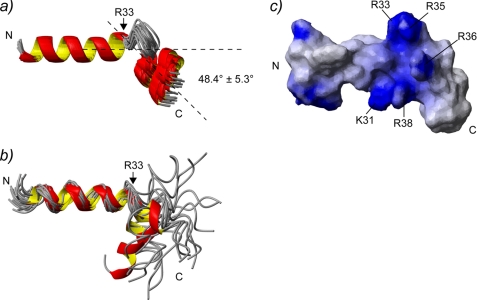
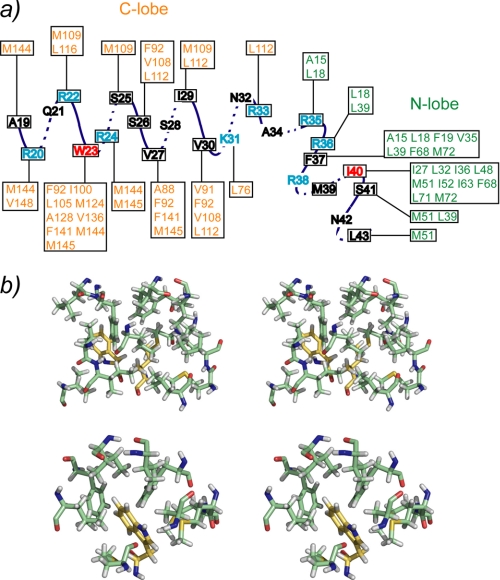
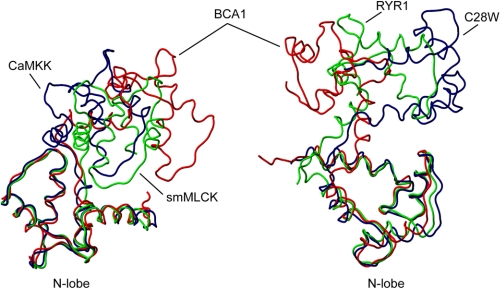
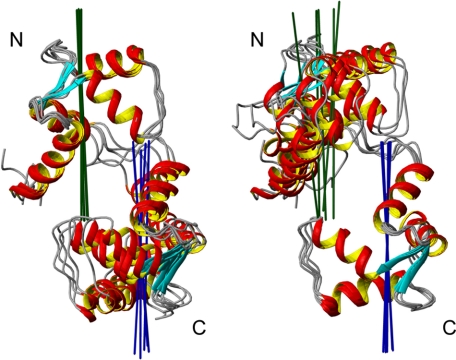
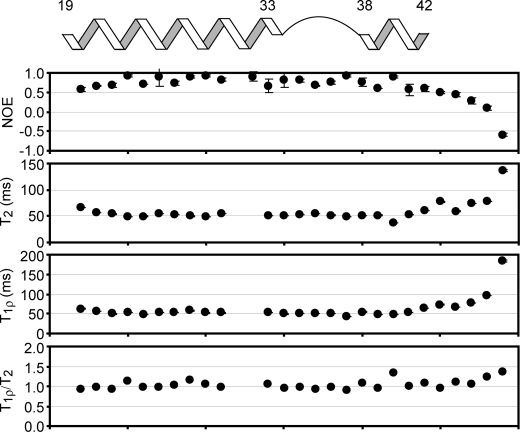
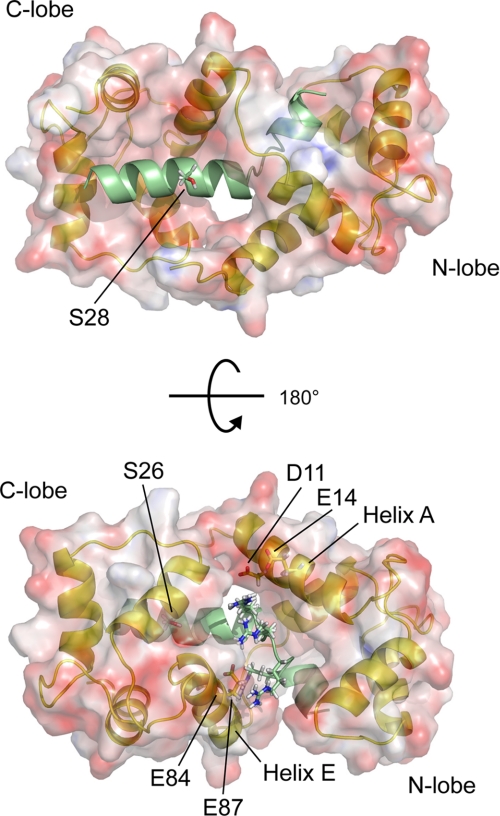
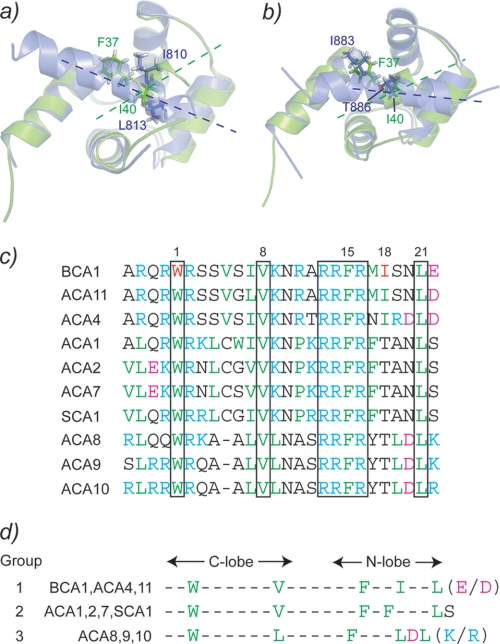
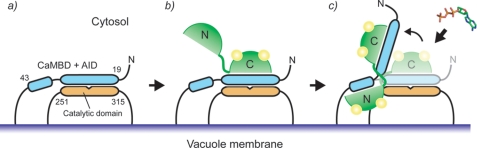
The type IIb class of plant Ca(2+)-ATPases contains a unique N-terminal extension that encompasses a calmodulin (CaM) binding domain and an auto-inhibitory domain. Binding of Ca(2+)-CaM to this region can release auto-inhibition and activates the calcium pump. Using multidimensional NMR spectroscopy, we have determined the solution structure of the complex of a plant CaM isoform with the CaM-binding domain of the well characterized Ca(2+)-ATPase BCA1 from cauliflower. The complex has a rather elongated structure in which the two lobes of CaM do not contact each other. The anchor residues Trp-23 and Ile-40 form a 1-8-18 interaction motif. Binding of Ca(2+)-CaM gives rise to the induction of two helical parts in this unique target peptide. The two helical portions are connected by a highly positively charged bend region, which represents a relatively fixed angle and positions the two lobes of CaM in an orientation that has not been seen before in any complex structure of calmodulin. The behavior of the complex was further characterized by heteronuclear NMR dynamics measurements of the isotope-labeled protein and peptide. These data suggest a unique calcium-driven activation mechanism for BCA1 and other plant Ca(2+)-ATPases that may also explain the action of calcium-CaM on some other target enzymes. Moreover, CaM activation of plant Ca(2+)-ATPases seems to occur in an organelle-specific manner.
Figures
References
-
- Reddy A. S. (2001) Plant Sci. 160, 381–404 - PubMed
-
- Lecourieux D., Ranjeva R., Pugin A. (2006) New Phytol. 171, 249–269 - PubMed
-
- Snedden W. A., Fromm H. (2001) New Phytol. 151, 35–66 - PubMed
-
- DeFalco T. A., Bender K. W., Snedden W. A. (2010) Biochem. J. 425, 27–40 - PubMed
-
- Zielinski R. E. (1998) Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 697–725 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
