

Closure of plasmodesmata in maize (Zea mays) at low temperature: a new mechanism for inhibition of photosynthesis
- PMID: 20880933
- PMCID: PMC2958785
- DOI: 10.1093/aob/mcq169
Closure of plasmodesmata in maize (Zea mays) at low temperature: a new mechanism for inhibition of photosynthesis
Abstract
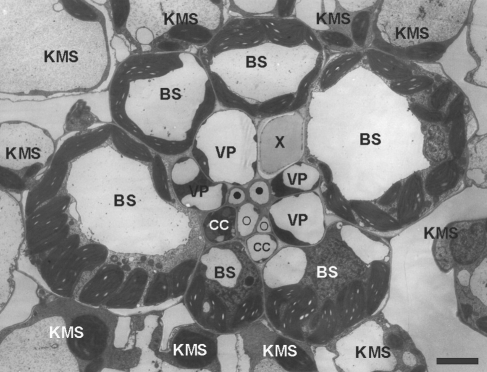
Background and aims: Photosynthesis is one of the processes most susceptible to low-temperature inhibition in maize, a tropical C4 crop not yet fully adapted to a temperate climate. C4 photosynthesis relies on symplasmic exchange of large amounts of photosynthetic intermediates between Kranz mesophyll (KMS) and bundle sheath (BS) cells. The aim of this study was to test the hypothesis that the slowing of maize photosynthesis at low temperature is related to ultrastructural changes in the plasmodesmata between KM and BS as well as BS and vascular parenchyma (VP) cells.
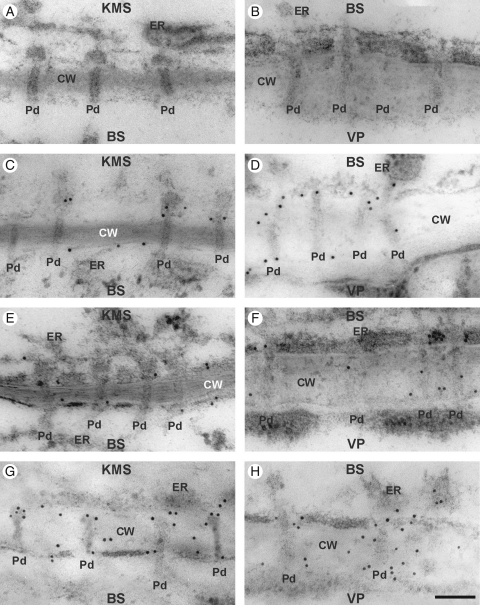
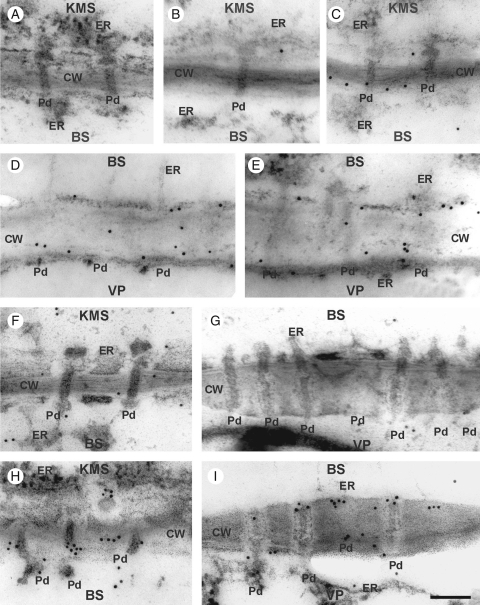
Methods: Chilling-tolerant (CT) KW 1074 and chilling-sensitive (CS) CM 109 maize (Zea mays) lines were studied. The effect of moderate chilling (14 °C) on the rate of photosynthesis, photosynthate transport kinetics, and the ultrastructure of plasmodesmata linking the KMS, BS and VP cells were analysed. Additionally, the accumulation of callose and calreticulin was studied by the immunogold method.
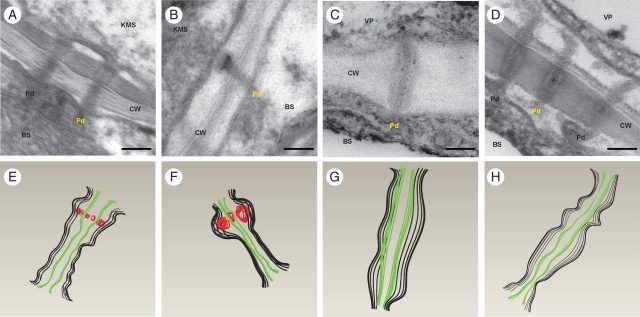
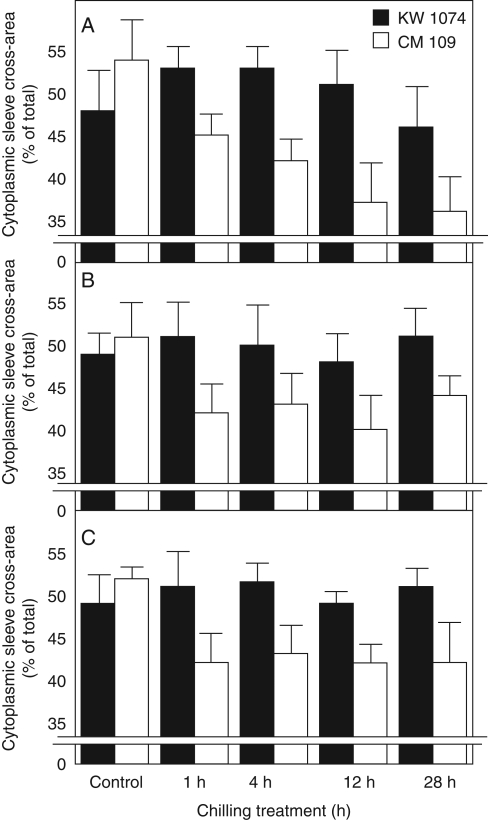
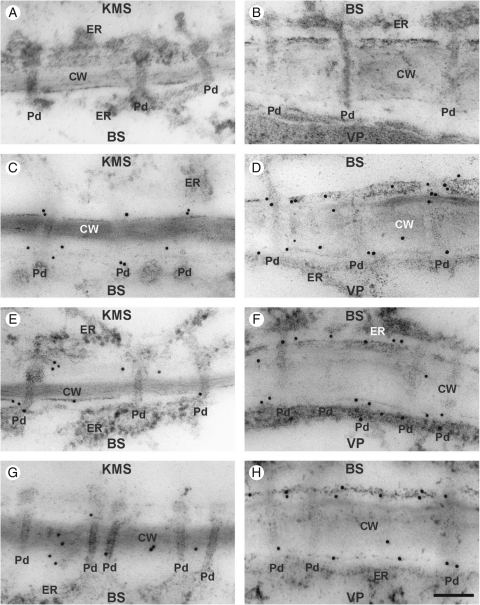
Key results: Chilling inhibited photosynthesis, photosynthate transfer to the phloem and photosynthate export from leaves in both lines. This inhibition was reversible upon cessation of chilling in the CT line but irreversible in the CS line. Simultaneously to physiological changes, chilling induced swelling of the sphincters of plasmodesmata linking KMS and BS cells and a decreased lumen of the cytoplasmic sleeve of plasmodesmata at the BS/VP interface in the CS line but not in the CT line. Accumulation of calreticulin, which occurred near the neck region of the closed plasmodesmata was observed after just 4 h of chilling and over-accumulation of callose at the KMS/BS and BS/VP interfaces occurred after 28 h of chilling.
Conclusions: Stronger chilling sensitivity of the CM 109 maize line compared with the KW 1074 line, shown by decreased photosynthesis and assimilate export from a leaf, is related to changes in the ultrastructure of leaf plasmodesmata at low temperature. The chain of reactions to chilling is likely to include calreticulin action resulting in rapid and efficient closure of the plasmodesmata at both KMS/BS and BS/VP interfaces. Callose deposition in a leaf was a secondary effect of chilling.
Figures
References
-
- Anisimov AV, Egorov AG. Plasmodesmata as a modulator of osmotic water fluxes in plants. Russian Journal of Plant Physiology. 2002;49:677–684.
-
- Aoki N, Hirose T, Takahashi S, Ono K, Ishimaru K, Ohsugi R. Molecular cloning and expression analysis of a gene for a sucrose transporter in maize (Zea mays L.) Plant and Cell Physiology. 1999;40:1072–1078. - PubMed
-
- Baluška F, Šamaj J, Napier R, Volkmann D. Maize calreticulin localizes preferentially to plasmodesmata in root apex. The Plant Journal. 1999;19:481–488. - PubMed
-
- Blackmann LM, Harper JDI, Overall RL. Localization of a centrin-like protein to higher plant plasmodesmata. European Journal of Cell Biology. 1999;78:297–304. - PubMed
-
- Botha CEJ, Cross RHM. Towards reconciliation of structure with function in plasmodesmata: who is gatekeeper? Micron. 2000;31:713–721. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
