

PP2B/calcineurin-mediated desensitization of TRPV1 does not require AKAP150
- PMID: 20883208
- PMCID: PMC3050517
- DOI: 10.1042/BJ20100936
PP2B/calcineurin-mediated desensitization of TRPV1 does not require AKAP150
Abstract
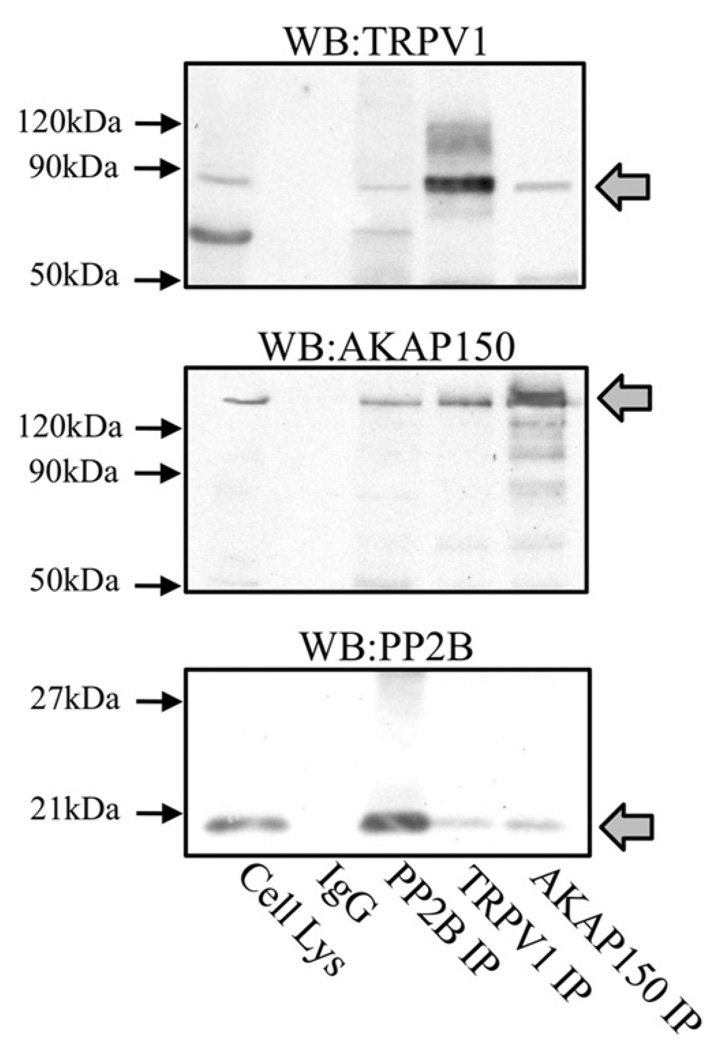
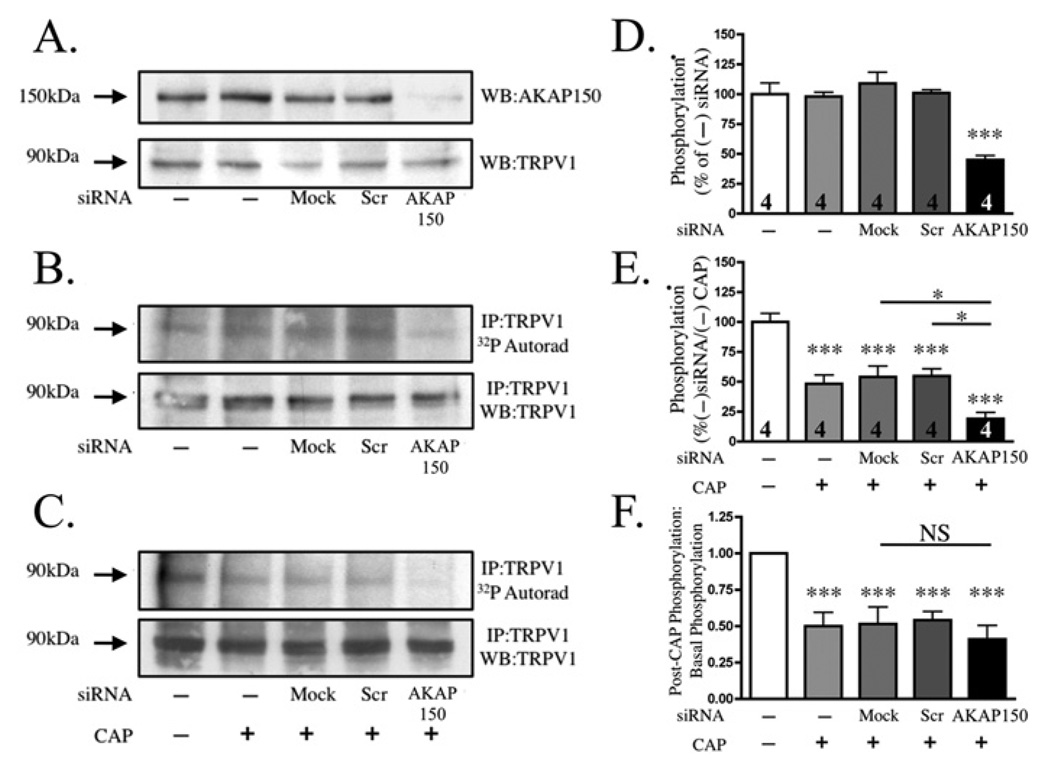
Activation of protein kinases and phosphatases at the plasma membrane often initiates agonist-dependent signalling events. In sensory neurons, AKAP150 (A-kinase-anchoring protein 150) orientates PKA (protein kinase A), PKC (protein kinase C) and the Ca2+/calmodulin-dependent PP2B (protein phosphatase 2B, also known as calcineurin) towards membrane-associated substrates. Recent evidence indicates that AKAP150-anchored PKA and PKC phosphorylate and sensitize the TRPV1 (transient receptor potential subfamily V type 1 channel, also known as the capsaicin receptor). In the present study, we explore the hypothesis that an AKAP150-associated pool of PP2B catalyses the dephosphorylation and desensitization of TRPV1. Biochemical, electrophysiological and cell-based experiments indicate that PP2B associates with AKAP150 and TRPV1 in cultured TG (trigeminal ganglia) neurons. Gene silencing of AKAP150 reduces basal phosphorylation of TRPV1. However, functional studies in neurons isolated from AKAP150-/- mice indicate that the anchoring protein is not required for pharmacological desensitization of TRPV1. Behavioural analysis of AKAP150-/- mice further support this notion, demonstrating that agonist-stimulated desensitization of TRPV1 is sensitive to PP2B inhibition and does not rely on AKAP150. These findings allow us to conclude that pharmacological desensitization of TRPV1 by PP2B may involve additional regulatory components.
Figures





References
-
- Wong W, Scott JD. AKAP signalling complexes: focal points in space and time. Nat. Rev. Mol. Cell Biol. 2004;5:959–970. - PubMed
-
- Klauck TM, Faux MC, Labudda K, Langeberg LK, Jaken S, Scott JD. Coordination of three signaling enzymes by AKAP79, a mammalian scaffold protein. Science. 1996;271:1589–1592. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous