

The DEAD-box RNA helicase-like Utp25 is an SSU processome component
- PMID: 20884785
- PMCID: PMC2957055
- DOI: 10.1261/rna.2359810
The DEAD-box RNA helicase-like Utp25 is an SSU processome component
Abstract
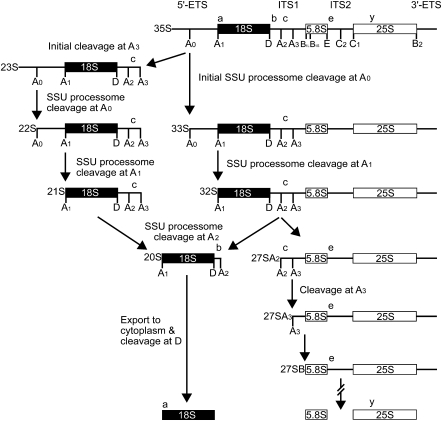
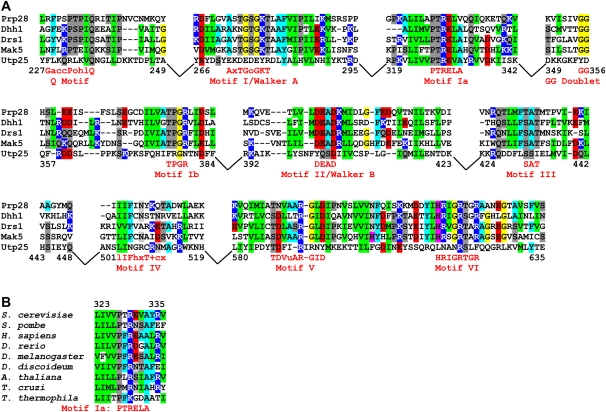
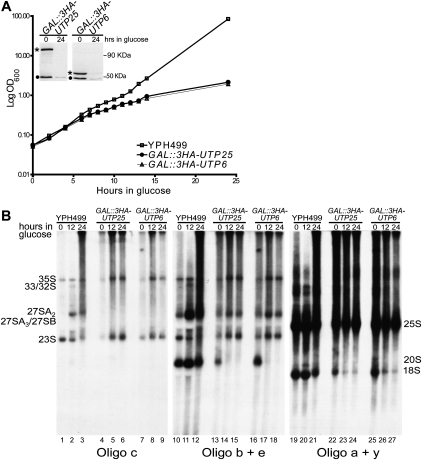
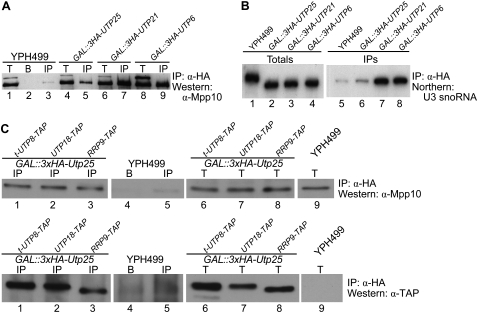
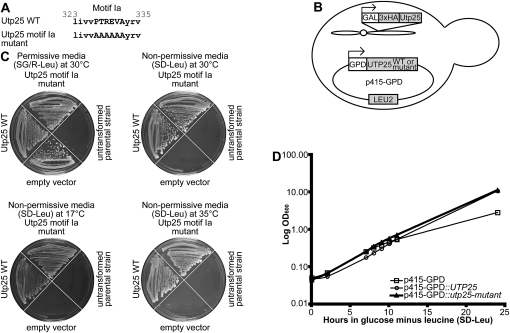
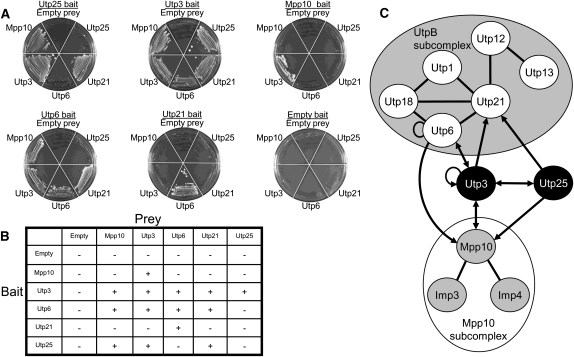
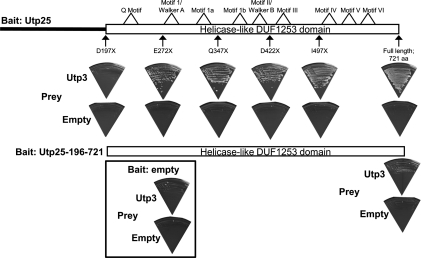
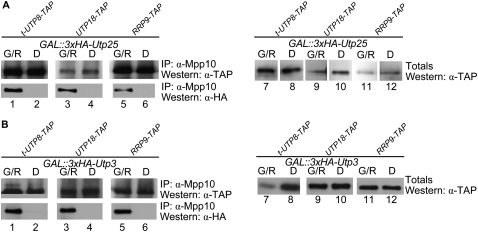
The SSU processome is a large ribonucleoprotein complex consisting of the U3 snoRNA and at least 43 proteins. A database search, initiated in an effort to discover additional SSU processome components, identified the uncharacterized, conserved and essential yeast nucleolar protein YIL091C/UTP25 as one such candidate. The C-terminal DUF1253 motif, a domain of unknown function, displays limited sequence similarity to DEAD-box RNA helicases. In the absence of the conserved DEAD-box sequence, motif Ia is the only clearly identifiable helicase element. Since the yeast homolog is nucleolar and interacts with components of the SSU processome, we examined its role in pre-rRNA processing. Genetic depletion of Utp25 resulted in slowed growth. Northern analysis of pre-rRNA revealed an 18S rRNA maturation defect at sites A₀, A₁, and A₂. Coimmunoprecipitation confirmed association with U3 snoRNA and with Mpp10, and with components of the t-Utp/UtpA, UtpB, and U3 snoRNP subcomplexes. Mutation of the conserved motif Ia residues resulted in no discernable temperature-sensitive or cold-sensitive growth defects, implying that this motif is dispensable for Utp25 function. A yeast two-hybrid screen of Utp25 against other SSU processome components revealed several interacting proteins, including Mpp10, Utp3, and Utp21, thereby identifying the first interactions among the different subcomplexes of the SSU processome. Furthermore, the DUF1253 domain is required and sufficient for the interaction of Utp25 with Utp3. Thus, Utp25 is a novel SSU processome component that, along with Utp3, forms the first identified interactions among the different SSU processome subcomplexes.
Figures
Similar articles
-
The small subunit processome in ribosome biogenesis—progress and prospects.Wiley Interdiscip Rev RNA. 2011 Jan-Feb;2(1):1-21. doi: 10.1002/wrna.57. Wiley Interdiscip Rev RNA. 2011. PMID: 21318072 Free PMC article. Review.
-
hUTP24 is essential for processing of the human rRNA precursor at site A1, but not at site A0.RNA Biol. 2015;12(9):1010-29. doi: 10.1080/15476286.2015.1073437. RNA Biol. 2015. PMID: 26237581 Free PMC article.
-
Utp14 interaction with the small subunit processome.RNA. 2018 Sep;24(9):1214-1228. doi: 10.1261/rna.066373.118. Epub 2018 Jun 20. RNA. 2018. PMID: 29925570 Free PMC article.
-
DEAD-box RNA helicase Dbp4 is required for small-subunit processome formation and function.Mol Cell Biol. 2015 Mar;35(5):816-30. doi: 10.1128/MCB.01348-14. Epub 2014 Dec 22. Mol Cell Biol. 2015. PMID: 25535329 Free PMC article.
-
The Ded1/DDX3 subfamily of DEAD-box RNA helicases.Crit Rev Biochem Mol Biol. 2014 Jul-Aug;49(4):343-60. doi: 10.3109/10409238.2014.931339. Crit Rev Biochem Mol Biol. 2014. PMID: 25039764 Review.
Cited by
-
Assembling a protein-protein interaction map of the SSU processome from existing datasets.PLoS One. 2011 Mar 10;6(3):e17701. doi: 10.1371/journal.pone.0017701. PLoS One. 2011. PMID: 21423703 Free PMC article.
-
The small subunit processome in ribosome biogenesis—progress and prospects.Wiley Interdiscip Rev RNA. 2011 Jan-Feb;2(1):1-21. doi: 10.1002/wrna.57. Wiley Interdiscip Rev RNA. 2011. PMID: 21318072 Free PMC article. Review.
-
Analysis of Hepatic Lentiviral Vector Transduction: Implications for Preclinical Studies and Clinical Gene Therapy Protocols.Viruses. 2025 Feb 17;17(2):276. doi: 10.3390/v17020276. Viruses. 2025. PMID: 40007031 Free PMC article.
-
Nop9 is a PUF-like protein that prevents premature cleavage to correctly process pre-18S rRNA.Nat Commun. 2016 Oct 11;7:13085. doi: 10.1038/ncomms13085. Nat Commun. 2016. PMID: 27725644 Free PMC article.
-
Cross-linking mass spectrometry discovers, evaluates, and corroborates structures and protein-protein interactions in the human cell.Proc Natl Acad Sci U S A. 2023 Apr 25;120(17):e2219418120. doi: 10.1073/pnas.2219418120. Epub 2023 Apr 18. Proc Natl Acad Sci U S A. 2023. PMID: 37071682 Free PMC article.
References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ 1990. Basic local alignment search tool. J Mol Biol 215: 403–410 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases