

Myogenin and class II HDACs control neurogenic muscle atrophy by inducing E3 ubiquitin ligases
- PMID: 20887891
- PMCID: PMC2982779
- DOI: 10.1016/j.cell.2010.09.004
Myogenin and class II HDACs control neurogenic muscle atrophy by inducing E3 ubiquitin ligases
Abstract
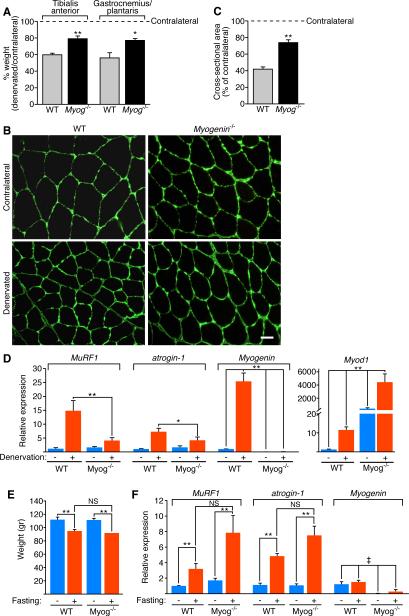
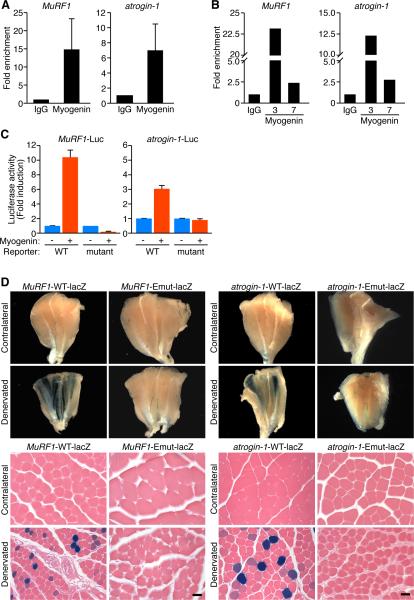
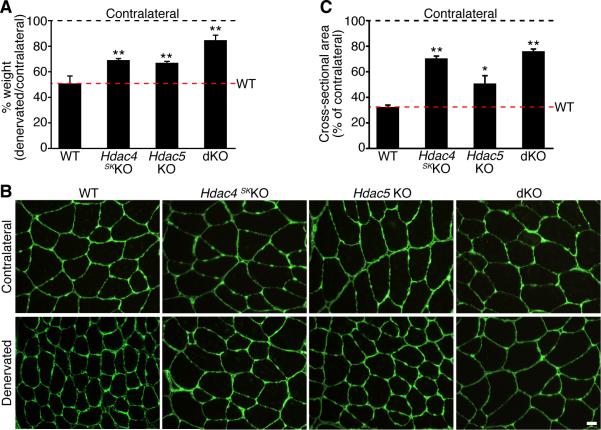
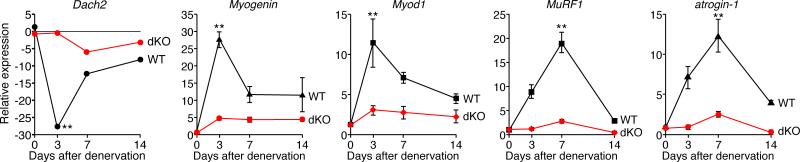
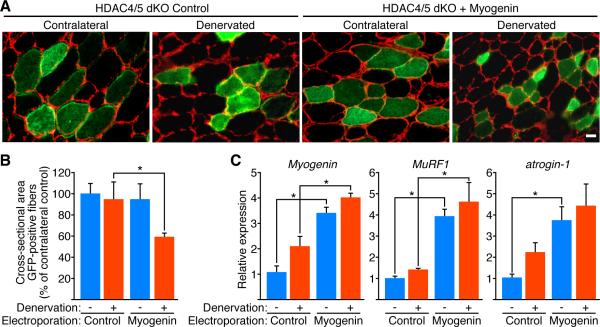
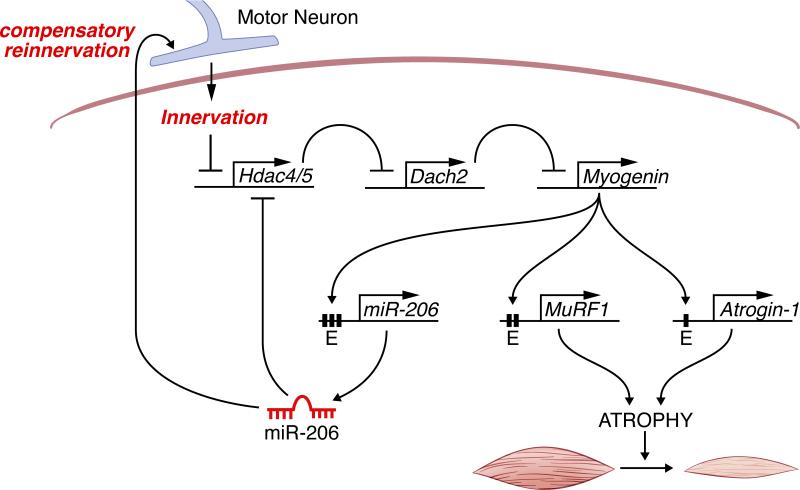
Maintenance of skeletal muscle structure and function requires innervation by motor neurons, such that denervation causes muscle atrophy. We show that myogenin, an essential regulator of muscle development, controls neurogenic atrophy. Myogenin is upregulated in skeletal muscle following denervation and regulates expression of the E3 ubiquitin ligases MuRF1 and atrogin-1, which promote muscle proteolysis and atrophy. Deletion of myogenin from adult mice diminishes expression of MuRF1 and atrogin-1 in denervated muscle and confers resistance to atrophy. Mice lacking histone deacetylases (HDACs) 4 and 5 in skeletal muscle fail to upregulate myogenin and also preserve muscle mass following denervation. Conversely, forced expression of myogenin in skeletal muscle of HDAC mutant mice restores muscle atrophy following denervation. Thus, myogenin plays a dual role as both a regulator of muscle development and an inducer of neurogenic atrophy. These findings reveal a specific pathway for muscle wasting and potential therapeutic targets for this disorder.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Attaix D, Combaret L, Bechet D, Taillandier D. Role of the ubiquitin-proteasome pathway in muscle atrophy in cachexia. Curr. Opin. Support Palliat. Care. 2008;2:262–266. - PubMed
-
- Attaix D, Ventadour S, Codran A, Bechet D, Taillandier D, Combaret L. The ubiquitin-proteasome system and skeletal muscle wasting. Essays Biochem. 2005;41:173–186. - PubMed
-
- Beehler BC, Sleph PG, Benmassaoud L, Grover GJ. Reduction of skeletal muscle atrophy by a proteasome inhibitor in a rat model of denervation. Exp. Biol. Med. 2006;231:335–341. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
