

Systematic protein location mapping reveals five principal chromatin types in Drosophila cells
- PMID: 20888037
- PMCID: PMC3119929
- DOI: 10.1016/j.cell.2010.09.009
Systematic protein location mapping reveals five principal chromatin types in Drosophila cells
Erratum in
- Cell. 2011 Apr 1;145(1):160
Abstract
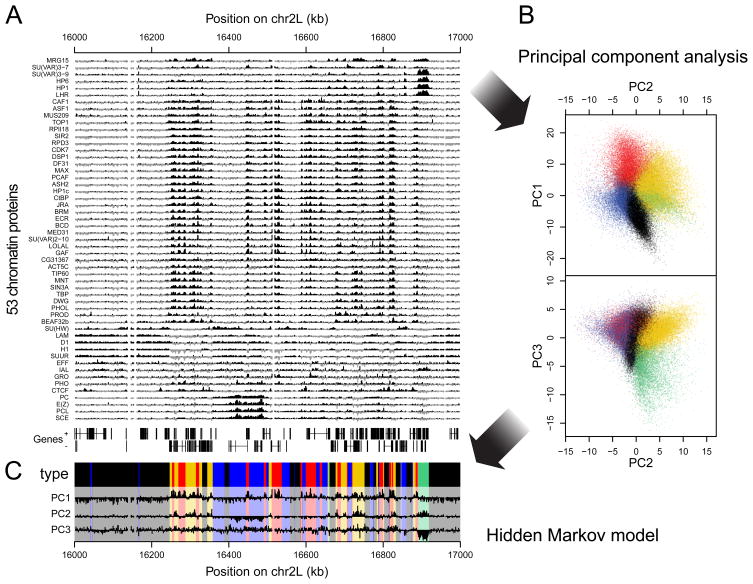
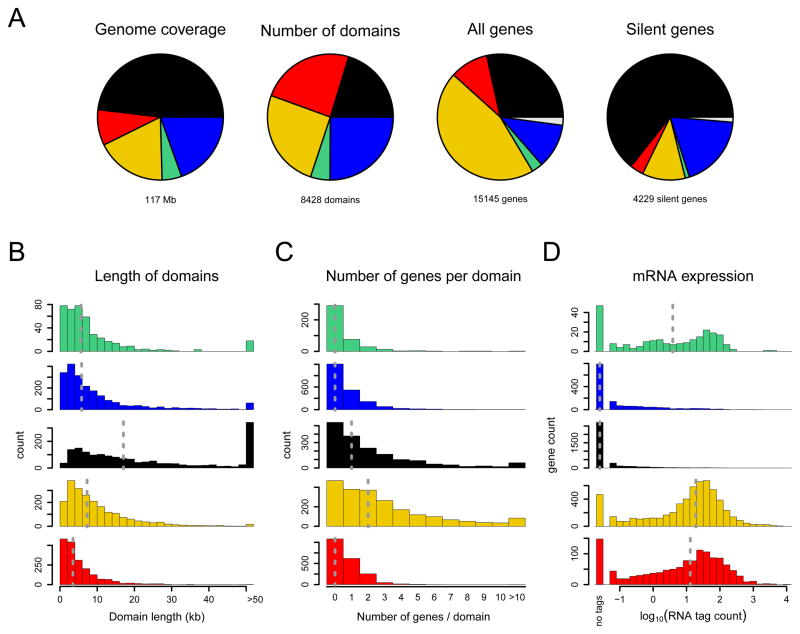
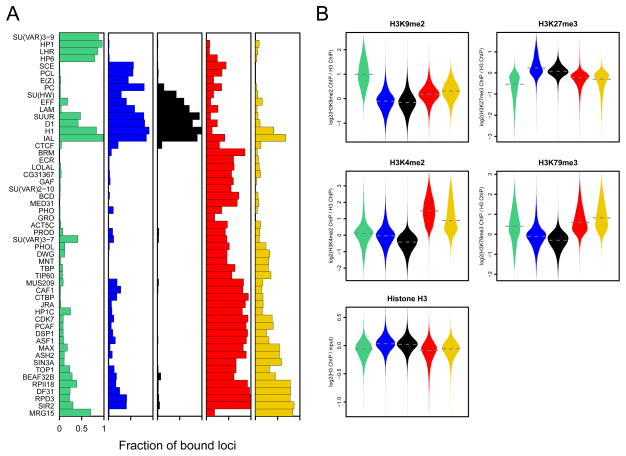
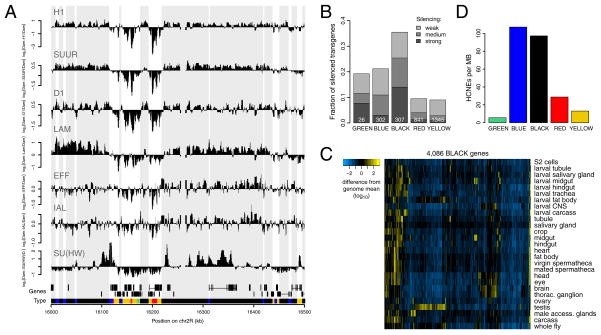
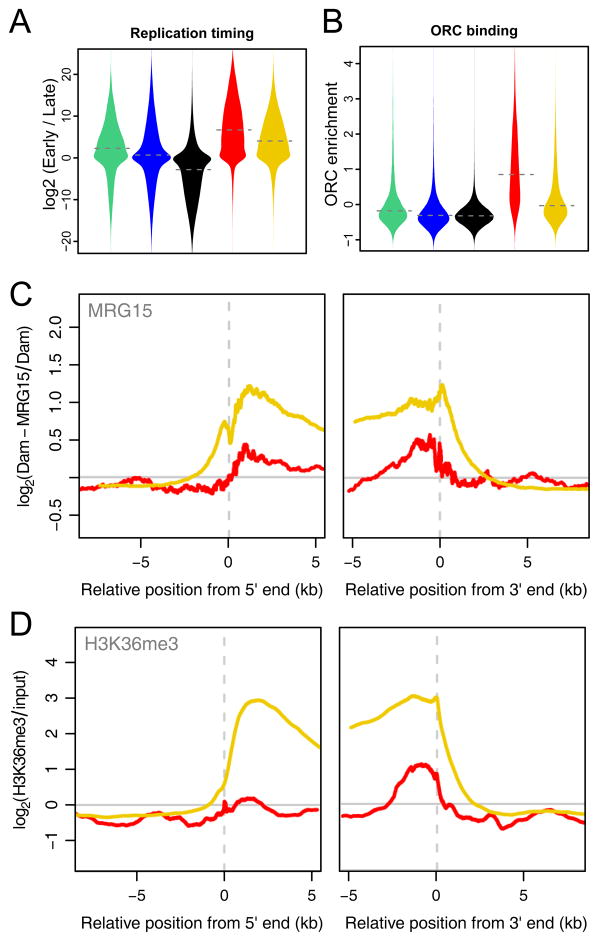
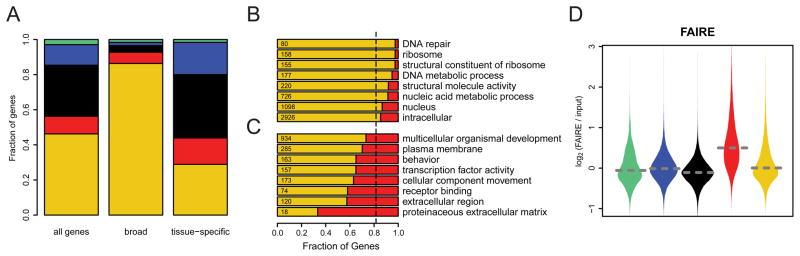
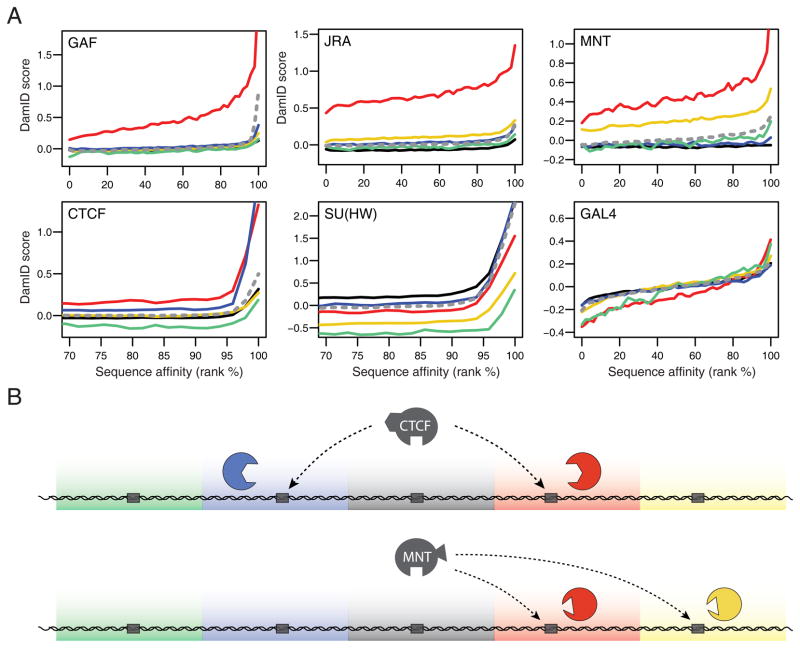
Chromatin is important for the regulation of transcription and other functions, yet the diversity of chromatin composition and the distribution along chromosomes are still poorly characterized. By integrative analysis of genome-wide binding maps of 53 broadly selected chromatin components in Drosophila cells, we show that the genome is segmented into five principal chromatin types that are defined by unique yet overlapping combinations of proteins and form domains that can extend over > 100 kb. We identify a repressive chromatin type that covers about half of the genome and lacks classic heterochromatin markers. Furthermore, transcriptionally active euchromatin consists of two types that differ in molecular organization and H3K36 methylation and regulate distinct classes of genes. Finally, we provide evidence that the different chromatin types help to target DNA-binding factors to specific genomic regions. These results provide a global view of chromatin diversity and domain organization in a metazoan cell.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Chromatin: Colour-coded classification.Nat Rev Genet. 2010 Nov;11(11):745. doi: 10.1038/nrg2887. Epub 2010 Oct 13. Nat Rev Genet. 2010. PMID: 20940735 No abstract available.
-
Chromatin in multicolor.Cell. 2010 Oct 15;143(2):183-4. doi: 10.1016/j.cell.2010.09.045. Cell. 2010. PMID: 20946976
References
-
- Bell O, Schwaiger M, Oakeley EJ, Lienert F, Beisel C, Stadler MB, Schubeler D. Accessibility of the Drosophila genome discriminates PcG repression, H4K16 acetylation and replication timing. Nat Struct Mol Biol. 2010;17:894–900. - PubMed
-
- Berger SL. The complex language of chromatin regulation during transcription. Nature. 2007;447:407–412. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
