

Studying bacterial transcriptomes using RNA-seq
- PMID: 20888288
- PMCID: PMC3025319
- DOI: 10.1016/j.mib.2010.09.009
Studying bacterial transcriptomes using RNA-seq
Abstract
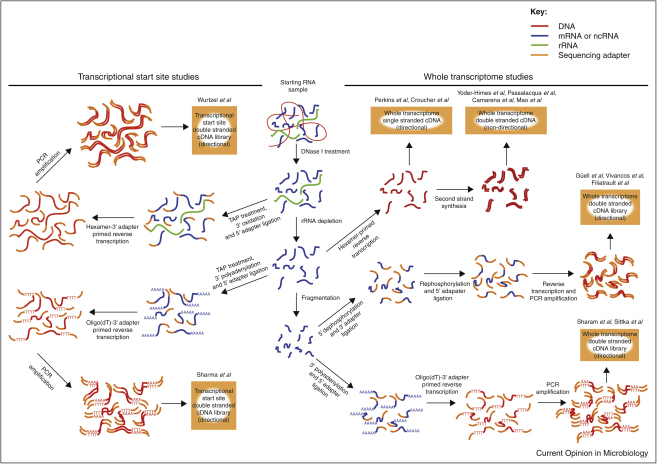
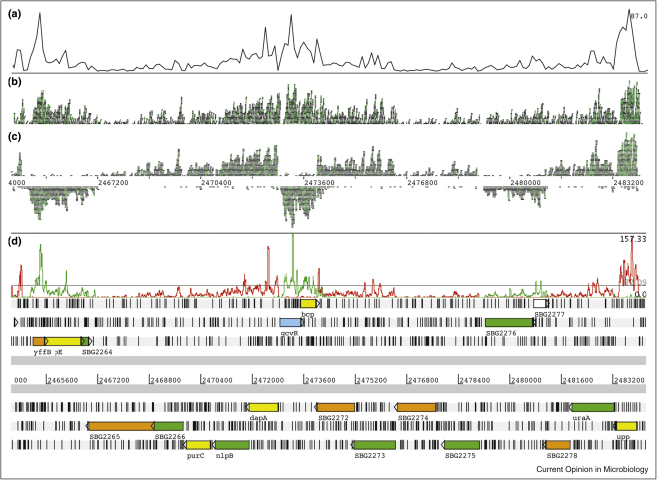
Genome-wide studies of bacterial gene expression are shifting from microarray technology to second generation sequencing platforms. RNA-seq has a number of advantages over hybridization-based techniques, such as annotation-independent detection of transcription, improved sensitivity and increased dynamic range. Early studies have uncovered a wealth of novel coding sequences and non-coding RNA, and are revealing a transcriptional landscape that increasingly mirrors that of eukaryotes. Already basic RNA-seq protocols have been improved and adapted to looking at particular aspects of RNA biology, often with an emphasis on non-coding RNAs, and further refinements to current techniques will improve our understanding of gene expression, and genome content, in the future.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Tettelin H., Riley D., Cattuto C., Medini D. Comparative genomics: the bacterial pan-genome. Curr Opin Microbiol. 2008;11:472–477. - PubMed
-
- Mortazavi A., Williams B.A., McCue K., Schaeffer L., Wold B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat Methods. 2008;5:621–628. - PubMed
-
- Oliver H.F., Orsi R.H., Ponnala L., Keich U., Wang W., Sun Q., Cartinhour S.W., Filiatrault M.J., Wiedmann M., Boor K.J. Deep RNA sequencing of L. monocytogenes reveals overlapping and extensive stationary phase and sigma B-dependent transcriptomes, including multiple highly transcribed noncoding RNAs. BMC Genomics. 2009;10:641. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
