

Evolutionary origin of a secondary structure: π-helices as cryptic but widespread insertional variations of α-helices that enhance protein functionality
- PMID: 20888342
- PMCID: PMC2981643
- DOI: 10.1016/j.jmb.2010.09.034
Evolutionary origin of a secondary structure: π-helices as cryptic but widespread insertional variations of α-helices that enhance protein functionality
Abstract
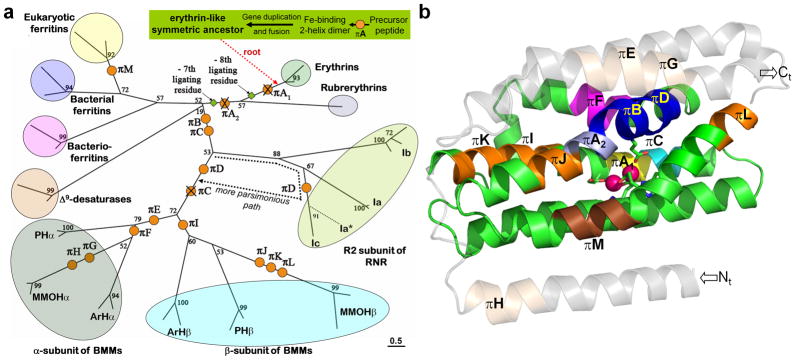
Formally annotated π-helices are rare in protein structures but have been correlated with functional sites. Here, we analyze protein structures to show that π-helices are the same as structures known as α-bulges, α-aneurisms, π-bulges, and looping outs, and are evolutionarily derived by the insertion of a single residue into an α-helix. This newly discovered evolutionary origin explains both why π-helices are cryptic, being rarely annotated despite occurring in 15% of known proteins, and why they tend to be associated with function. An analysis of π-helices in the diverse ferritin-like superfamily illustrates their tendency to be conserved in protein families and identifies a putative π-helix-containing primordial precursor, a "missing link" intermediary form of the ribonucleotide reductase family, vestigial π-helices, and a novel function for π-helices that we term a "peristaltic-like shift." This new understanding of π-helices paves the way for this generally overlooked motif to become a noteworthy feature that will aid in tracing the evolution of many protein families, guide investigations of protein and π-helix functionality, and contribute additional tools to the protein engineering toolkit.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures








References
-
- Worth CL, Gong S, Blundell TL. Structural and functional constraints in the evolution of protein families. Nat Rev Mol Cell Biol. 2009;10:709–20. - PubMed
-
- Low BW, Baybutt RB. The π-helix- A Hydrogen Bonded Configuration of the Polypeptide Chain. J Am Chem Soc. 1952;74:5806–5807.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
