

Colocalization of amanitin and a candidate toxin-processing prolyl oligopeptidase in Amanita basidiocarps
- PMID: 20889720
- PMCID: PMC3008271
- DOI: 10.1128/EC.00161-10
Colocalization of amanitin and a candidate toxin-processing prolyl oligopeptidase in Amanita basidiocarps
Abstract
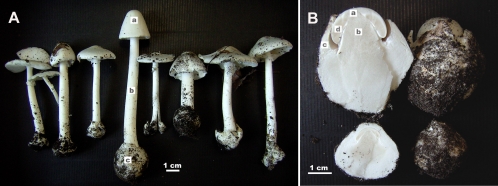
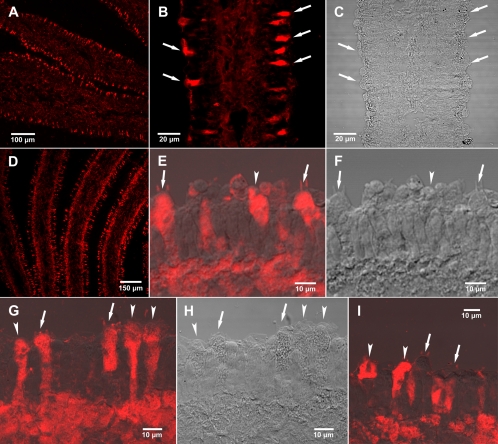
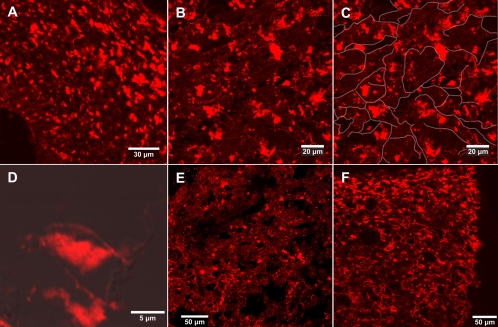
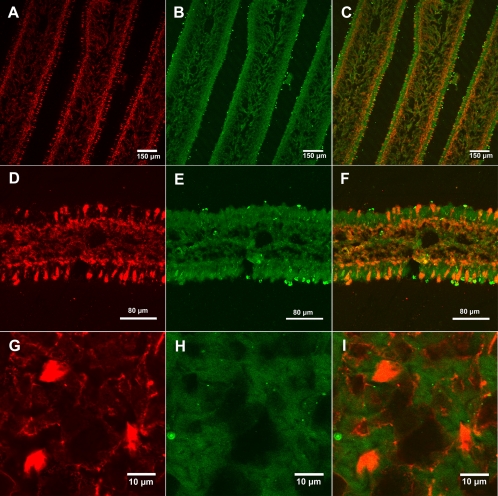
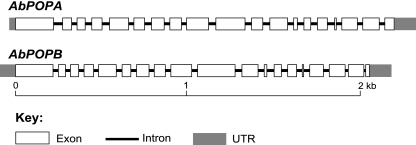
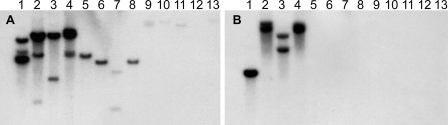
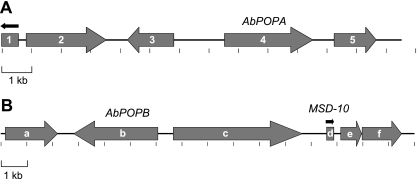
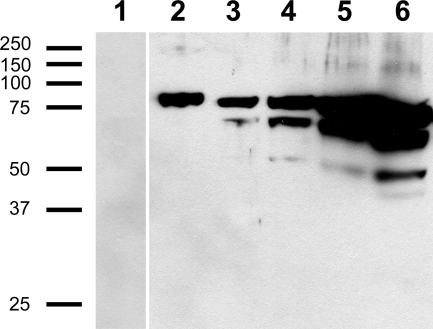
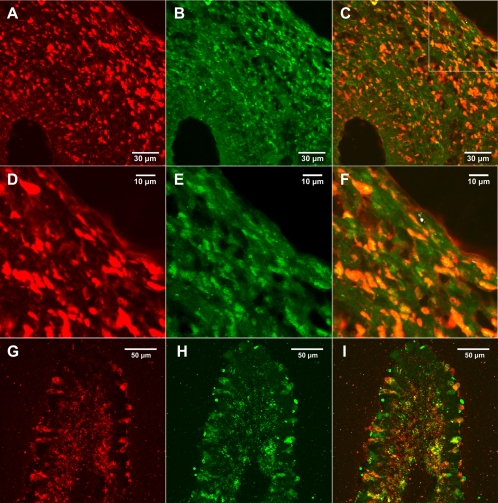
Fungi in the basidiomycetous genus Amanita owe their high mammalian toxicity to the bicyclic octapeptide amatoxins such as α-amanitin. Amatoxins and the related phallotoxins (such as the heptapeptide phalloidin) are encoded by members of the "MSDIN" gene family and are synthesized on ribosomes as short (34- to 35-amino-acid) proproteins. Antiamanitin antibodies and confocal microscopy were used to determine the cellular and subcellular localizations of amanitin accumulation in basidiocarps (mushrooms) of the Eastern North American destroying angel (Amanita bisporigera). Consistent with previous studies, amanitin is present throughout the basidiocarp (stipe, pileus, lamellae, trama, and universal veil), but it is present in only a subset of cells within these tissues. Restriction of amanitin to certain cells is especially marked in the hymenium. Several lines of evidence implicate a specific prolyl oligopeptidase, A. bisporigera POPB (AbPOPB), in the initial processing of the amanitin and phallotoxin proproteins. The gene for AbPOPB is restricted taxonomically to the amatoxin-producing species of Amanita and is clustered in the genome with at least one expressed member of the MSDIN gene family. Immunologically, amanitin and AbPOPB show a high degree of colocalization, indicating that toxin biosynthesis and accumulation occur in the same cells and possibly in the same subcellular compartments.
Figures
Similar articles
-
Diversity of MSDIN family members in amanitin-producing mushrooms and the phylogeny of the MSDIN and prolyl oligopeptidase genes.BMC Genomics. 2020 Jun 26;21(1):440. doi: 10.1186/s12864-020-06857-8. BMC Genomics. 2020. PMID: 32590929 Free PMC article.
-
Gene family encoding the major toxins of lethal Amanita mushrooms.Proc Natl Acad Sci U S A. 2007 Nov 27;104(48):19097-101. doi: 10.1073/pnas.0707340104. Epub 2007 Nov 19. Proc Natl Acad Sci U S A. 2007. PMID: 18025465 Free PMC article.
-
Ribosomal biosynthesis of α-amanitin in Galerina marginata.Fungal Genet Biol. 2012 Feb;49(2):123-9. doi: 10.1016/j.fgb.2011.12.005. Epub 2011 Dec 21. Fungal Genet Biol. 2012. PMID: 22202811 Free PMC article.
-
Ribosomal biosynthesis of the cyclic peptide toxins of Amanita mushrooms.Biopolymers. 2010;94(5):659-64. doi: 10.1002/bip.21416. Biopolymers. 2010. PMID: 20564017 Free PMC article. Review.
-
Mechanism and treatment of α-amanitin poisoning.Arch Toxicol. 2023 Jan;97(1):121-131. doi: 10.1007/s00204-022-03396-x. Epub 2022 Oct 21. Arch Toxicol. 2023. PMID: 36271256 Review.
Cited by
-
Exploring fungal RiPPs from the perspective of chemical ecology.Fungal Biol Biotechnol. 2022 Jun 25;9(1):12. doi: 10.1186/s40694-022-00144-9. Fungal Biol Biotechnol. 2022. PMID: 35752794 Free PMC article. Review.
-
Circular proteins from plants and fungi.J Biol Chem. 2012 Aug 3;287(32):27001-6. doi: 10.1074/jbc.R111.300129. Epub 2012 Jun 14. J Biol Chem. 2012. PMID: 22700984 Free PMC article. Review.
-
Genome of lethal Lepiota venenata and insights into the evolution of toxin-biosynthetic genes.BMC Genomics. 2019 Mar 8;20(1):198. doi: 10.1186/s12864-019-5575-7. BMC Genomics. 2019. PMID: 30849934 Free PMC article.
-
Phylogenomic Analyses of Non-Dikarya Fungi Supports Horizontal Gene Transfer Driving Diversification of Secondary Metabolism in the Amphibian Gastrointestinal Symbiont, Basidiobolus.G3 (Bethesda). 2020 Sep 2;10(9):3417-3433. doi: 10.1534/g3.120.401516. G3 (Bethesda). 2020. PMID: 32727924 Free PMC article.
-
Subcellular localization of fungal specialized metabolites.Fungal Biol Biotechnol. 2022 May 25;9(1):11. doi: 10.1186/s40694-022-00140-z. Fungal Biol Biotechnol. 2022. PMID: 35614515 Free PMC article. Review.
References
-
- Bastos I. M., Motta F. N., Charneau S., Santana J. M., Dubost L., Augustyns K., Grellier P. 2010. Prolyl oligopeptidase of Trypanosoma brucei hydrolyzes native collagen, peptide hormones and is active in the plasma of infected mice. Microbes Infect. 12:457–466 - PubMed
-
- Bresinsky A., Besl H. 1990. A colour atlas of poisonous fungi: a handbook for pharmacists, doctors and biologists, p. 295 Wolfe, Würzburg, Germany
-
- Clémençon H. 2004. Cytology and plectology of the Hymenomycetes, p. 24–50 J. Cramer, Berlin, Germany
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
