

Characterization of a novel riboswitch-regulated lysine transporter in Aggregatibacter actinomycetemcomitans
- PMID: 20889741
- PMCID: PMC2981213
- DOI: 10.1128/JB.00935-10
Characterization of a novel riboswitch-regulated lysine transporter in Aggregatibacter actinomycetemcomitans
Abstract
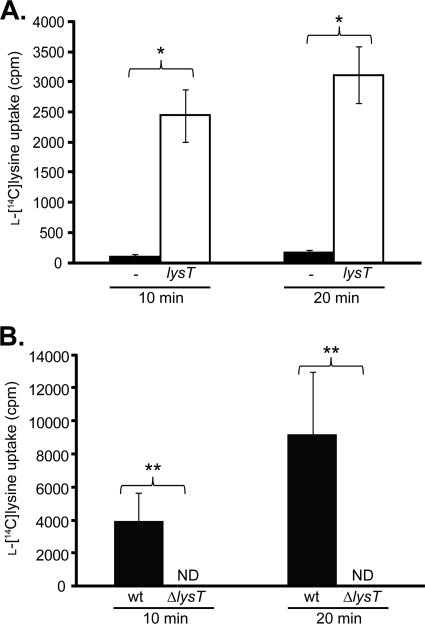
Aggregatibacter actinomycetemcomitans is an opportunistic pathogen that resides primarily in the mammalian oral cavity. In this environment, A. actinomycetemcomitans faces numerous host- and microbe-derived stresses, including intense competition for nutrients and exposure to the host immune system. While it is clear that A. actinomycetemcomitans responds to precise cues that allow it to adapt and proliferate in the presence of these stresses, little is currently known about the regulatory mechanisms that underlie these responses. Many bacteria use noncoding regulatory RNAs (ncRNAs) to rapidly alter gene expression in response to environmental stresses. Although no ncRNAs have been reported in A. actinomycetemcomitans, we propose that they are likely important for colonization and persistence in the oral cavity. Using a bioinformatic and experimental approach, we identified three putative metabolite-sensing riboswitches and nine small regulatory RNAs (sRNAs) in A. actinomycetemcomitans during planktonic and biofilm growth. Molecular characterization of one of the riboswitches revealed that it is a lysine riboswitch and that its target gene, lysT, encodes a novel lysine-specific transporter. Finally, we demonstrated that lysT and the lysT lysine riboswitch are conserved in over 40 bacterial species, including the phylogenetically related pathogen Haemophilus influenzae.
Figures






Similar articles
-
Comparative genomics and phylogenomic analyses of lysine riboswitch distributions in bacteria.PLoS One. 2017 Sep 5;12(9):e0184314. doi: 10.1371/journal.pone.0184314. eCollection 2017. PLoS One. 2017. PMID: 28873470 Free PMC article.
-
Probing bacterial metabolism during infection using high-resolution transcriptomics.J Bacteriol. 2013 Nov;195(22):4991-8. doi: 10.1128/JB.00875-13. Epub 2013 Aug 23. J Bacteriol. 2013. PMID: 23974023 Free PMC article.
-
QseBC, a two-component bacterial adrenergic receptor and global regulator of virulence in Enterobacteriaceae and Pasteurellaceae.Mol Oral Microbiol. 2016 Oct;31(5):379-97. doi: 10.1111/omi.12138. Epub 2015 Nov 20. Mol Oral Microbiol. 2016. PMID: 26426681 Free PMC article. Review.
-
Structural insights into amino acid binding and gene control by a lysine riboswitch.Nature. 2008 Oct 30;455(7217):1263-7. doi: 10.1038/nature07326. Epub 2008 Sep 10. Nature. 2008. PMID: 18784651 Free PMC article.
-
Mechanisms by Which Small RNAs Affect Bacterial Activity.J Dent Res. 2019 Nov;98(12):1315-1323. doi: 10.1177/0022034519876898. Epub 2019 Sep 23. J Dent Res. 2019. PMID: 31547763 Review.
Cited by
-
An in vitro model for studies of attenuation of antibiotic-inhibited growth of Aggregatibacter actinomycetemcomitans Y4 by polyamines.Mol Oral Microbiol. 2021 Dec;36(6):308-315. doi: 10.1111/omi.12353. Epub 2021 Sep 21. Mol Oral Microbiol. 2021. PMID: 34486245 Free PMC article.
-
Analysis of lysine recognition and specificity of the Bacillus subtilis L box riboswitch.Nucleic Acids Res. 2012 Jul;40(12):5706-17. doi: 10.1093/nar/gks212. Epub 2012 Mar 13. Nucleic Acids Res. 2012. PMID: 22416067 Free PMC article.
-
Small regulatory RNAs of oral streptococci and periodontal bacteria.Jpn Dent Sci Rev. 2021 Nov;57:209-216. doi: 10.1016/j.jdsr.2021.09.004. Epub 2021 Oct 23. Jpn Dent Sci Rev. 2021. PMID: 34745393 Free PMC article. Review.
-
Characterization of a transcriptional TPP riboswitch in the human pathogen Neisseria meningitidis.RNA Biol. 2020 May;17(5):718-730. doi: 10.1080/15476286.2020.1727188. Epub 2020 Feb 20. RNA Biol. 2020. PMID: 32079473 Free PMC article.
-
Identification of a glutathione transporter in A. actinomycetemcomitans.Microbiol Spectr. 2024 Jan 11;12(1):e0351123. doi: 10.1128/spectrum.03511-23. Epub 2023 Dec 5. Microbiol Spectr. 2024. PMID: 38051055 Free PMC article.
References
-
- Abe, N., A. Baba, R. Takii, K. Nakayama, A. Kamaguchi, Y. Shibata, Y. Abiko, K. Okamoto, T. Kadowaki, and K. Yamamoto. 2004. Roles of Arg- and Lys-gingipains in coaggregation of Porphyromonas gingivalis: identification of its responsible molecules in translation products of rgpA, kgp, and hagA genes. Biol. Chem. 385:1041-1047. - PubMed
-
- Ausubel, F. M. 2002. Short protocols in molecular biology: a compendium of methods from Current Protocols in Molecular Biology, 5th ed. Wiley, New York, NY.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
