

HU protein affects transcription of surface polysaccharide synthesis genes in Porphyromonas gingivalis
- PMID: 20889748
- PMCID: PMC2981211
- DOI: 10.1128/JB.00106-10
HU protein affects transcription of surface polysaccharide synthesis genes in Porphyromonas gingivalis
Abstract
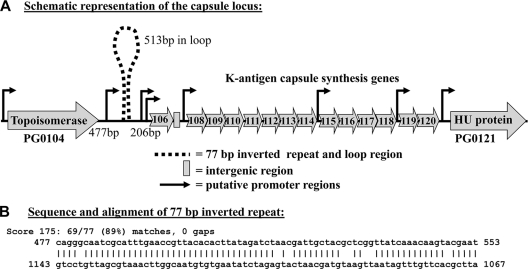
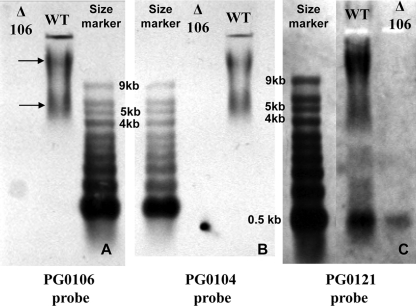
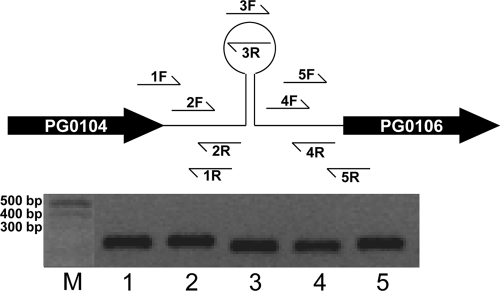
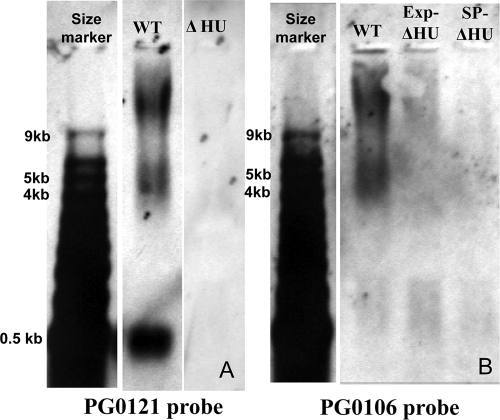
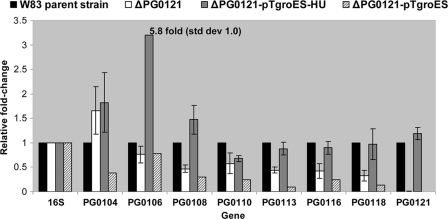
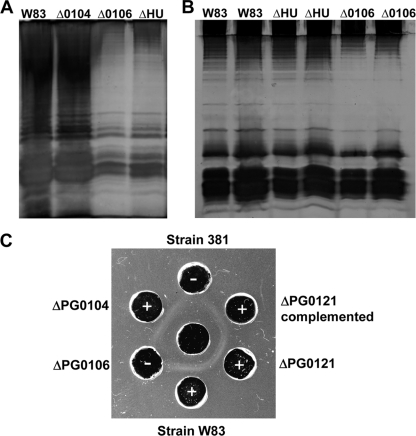
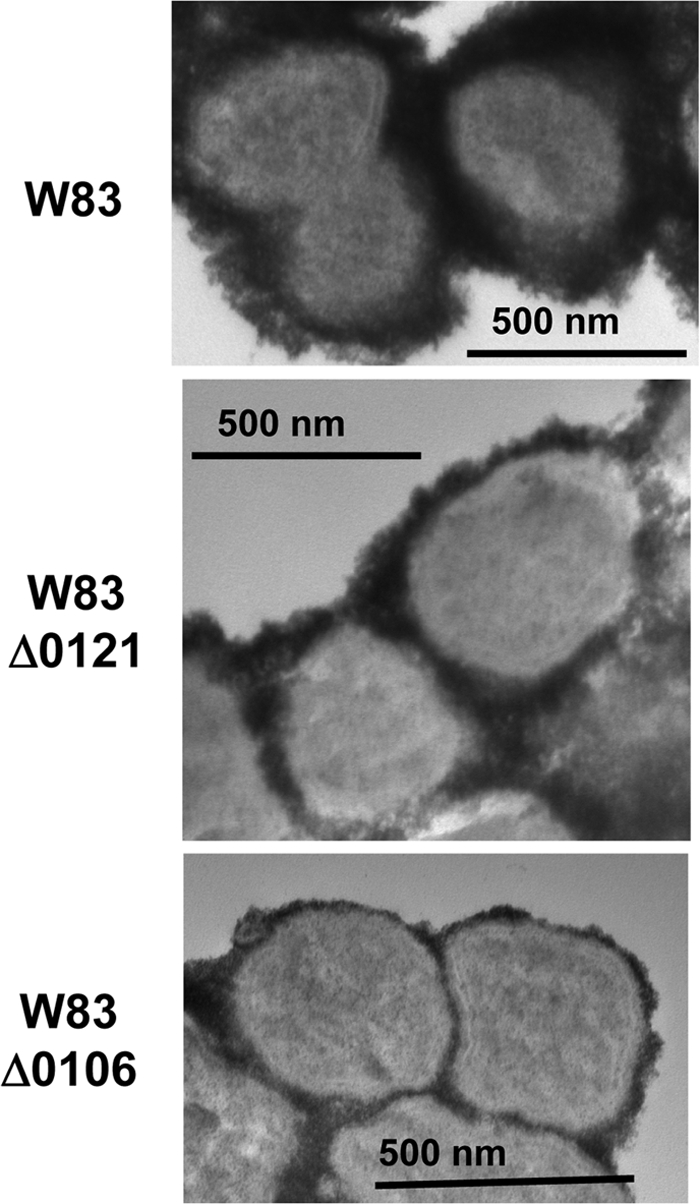
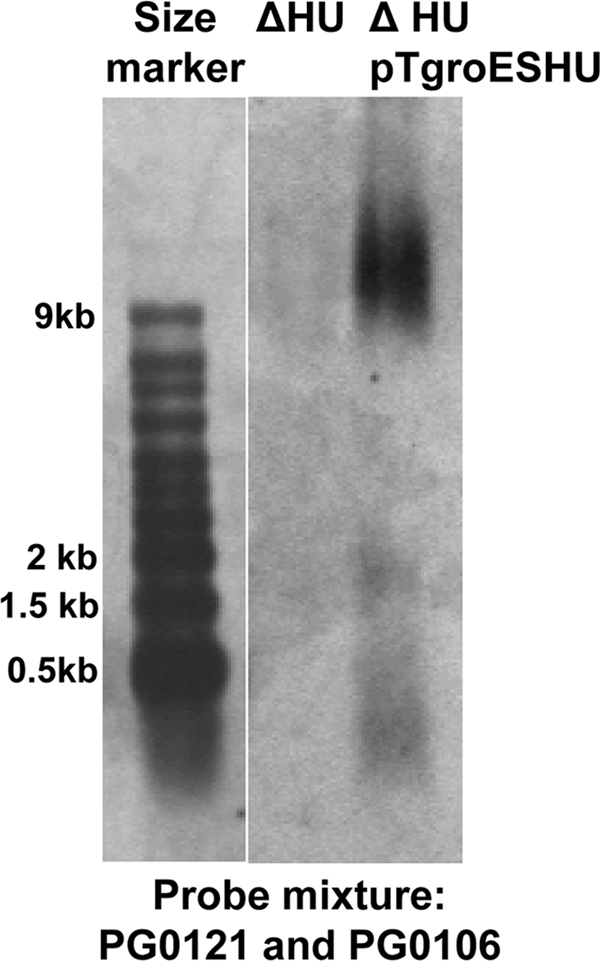
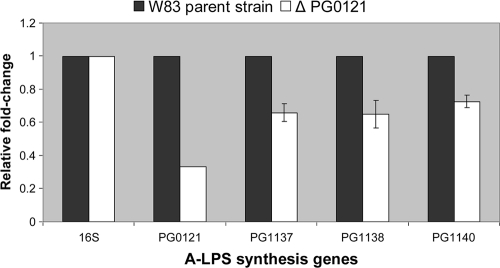
K-antigen capsule synthesis is an important virulence determinant of the oral anaerobe Porphyromonas gingivalis. We previously reported that the locus required for synthesis of this surface polysaccharide in strain W83 (TIGR identification PG0106 to PG0120) is transcribed as a large (∼16.7-kb) polycistronic message. Through sequence analysis, we have now identified a 77-bp inverted repeat located upstream (206 bp) of the start codon of PG0106 that is capable of forming a large hairpin structure. Further sequence analysis just upstream and downstream of the capsule synthesis genes revealed the presence of two genes oriented in the same direction as the operon that are predicted to encode DNA binding proteins: PG0104, which is highly similar (57%) to DNA topoisomerase III, and PG0121, which has high similarity (72%) to DNA binding protein HU (β-subunit). In this report, we show that these two genes, as well as the 77-bp inverted repeat region, are cotranscribed with the capsule synthesis genes, resulting in a large transcript that is ∼19.4 kb (based on annotation). We also show that a PG0121 recombinant protein is a nonspecific DNA binding protein with strong affinity to the hairpin structure, in vitro, and that transcript levels of the capsule synthesis genes are downregulated in a PG0121 deletion mutant. Furthermore, we show that this decrease in transcript levels corresponds to a decrease in the amount of polysaccharide produced. Interestingly, expression analysis of another polysaccharide synthesis locus (PG1136 to PG1143) encoding genes involved in synthesis of a surface-associated phosphorylated branched mannan (APS) indicated that this locus is also downregulated in the PG0121 mutant. Altogether our data indicate that HU protein modulates expression of surface polysaccharides in P. gingivalis strain W83.
Figures

Similar articles
-
A biochemical analysis of the interaction of Porphyromonas gingivalis HU PG0121 protein with DNA.PLoS One. 2014 Mar 28;9(3):e93266. doi: 10.1371/journal.pone.0093266. eCollection 2014. PLoS One. 2014. PMID: 24681691 Free PMC article.
-
Deletion of a 77-base-pair inverted repeat element alters the synthesis of surface polysaccharides in Porphyromonas gingivalis.J Bacteriol. 2015 Apr;197(7):1208-20. doi: 10.1128/JB.02589-14. Epub 2015 Jan 26. J Bacteriol. 2015. PMID: 25622614 Free PMC article.
-
The nucleoid-associated protein HUβ affects global gene expression in Porphyromonas gingivalis.Microbiology (Reading). 2013 Feb;159(Pt 2):219-229. doi: 10.1099/mic.0.061002-0. Epub 2012 Nov 22. Microbiology (Reading). 2013. PMID: 23175503 Free PMC article.
-
A Novel Regulation of K-antigen Capsule Synthesis in Porphyromonas gingivalis Is Driven by the Response Regulator PG0720-Directed Antisense RNA.Front Oral Health. 2021 Jul 1;2:701659. doi: 10.3389/froh.2021.701659. eCollection 2021. Front Oral Health. 2021. PMID: 35048039 Free PMC article.
-
Identification and characterization of the capsular polysaccharide (K-antigen) locus of Porphyromonas gingivalis.Infect Immun. 2006 Jan;74(1):449-60. doi: 10.1128/IAI.74.1.449-460.2006. Infect Immun. 2006. PMID: 16369001 Free PMC article.
Cited by
-
Genes Contributing to Porphyromonas gingivalis Fitness in Abscess and Epithelial Cell Colonization Environments.Front Cell Infect Microbiol. 2017 Aug 28;7:378. doi: 10.3389/fcimb.2017.00378. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28900609 Free PMC article.
-
DNA-Binding Protein HU Coordinates Pathogenicity in Vibrio parahaemolyticus.J Bacteriol. 2015 Sep;197(18):2958-64. doi: 10.1128/JB.00306-15. Epub 2015 Jul 6. J Bacteriol. 2015. PMID: 26148713 Free PMC article.
-
A biochemical analysis of the interaction of Porphyromonas gingivalis HU PG0121 protein with DNA.PLoS One. 2014 Mar 28;9(3):e93266. doi: 10.1371/journal.pone.0093266. eCollection 2014. PLoS One. 2014. PMID: 24681691 Free PMC article.
-
Microbial signatures in human periodontal disease: a metatranscriptome meta-analysis.Front Microbiol. 2024 Apr 9;15:1383404. doi: 10.3389/fmicb.2024.1383404. eCollection 2024. Front Microbiol. 2024. PMID: 38659984 Free PMC article.
-
The dental plaque biofilm matrix.Periodontol 2000. 2021 Jun;86(1):32-56. doi: 10.1111/prd.12361. Epub 2021 Mar 10. Periodontol 2000. 2021. PMID: 33690911 Free PMC article. Review.
References
-
- Aduse-Opoku, J., J. M. Slaney, A. Hashim, A. Gallagher, R. P. Gallagher, M. Rangarajan, K. Boutaga, M. L. Laine, A. J. Van Winkelhoff, and M. A. Curtis. 2006. Identification and characterization of the capsular polysaccharide (K-antigen) locus of Porphyromonas gingivalis. Infect. Immun. 74:449-460. - PMC - PubMed
-
- Balandina, A., L. Claret, R. Hengge-Aronis, and J. Rouviere-Yaniv. 2001. The Escherichia coli histone-like protein HU regulates rpoS translation. Mol. Microbiol. 39:1069-1079. - PubMed
-
- Balandina, A., D. Kamashev, and J. Rouviere-Yaniv. 2002. The bacterial histone-like protein HU specifically recognizes similar structures in all nucleic acids. DNA, RNA, and their hybrids. J. Biol. Chem. 277:27622-27628. - PubMed
-
- Bensaid, A., A. Almeida, K. Drlica, and J. Rouviere-Yaniv. 1996. Cross-talk between topoisomerase I and HU in Escherichia coli. J. Mol. Biol. 256:292-300. - PubMed
-
- Bonnefoy, E., M. Takahashi, and J. R. Yaniv. 1994. DNA-binding parameters of the HU protein of Escherichia coli to cruciform DNA. J. Mol. Biol. 242:116-129. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
