

Categorical speech representation in human superior temporal gyrus
- PMID: 20890293
- PMCID: PMC2967728
- DOI: 10.1038/nn.2641
Categorical speech representation in human superior temporal gyrus
Abstract
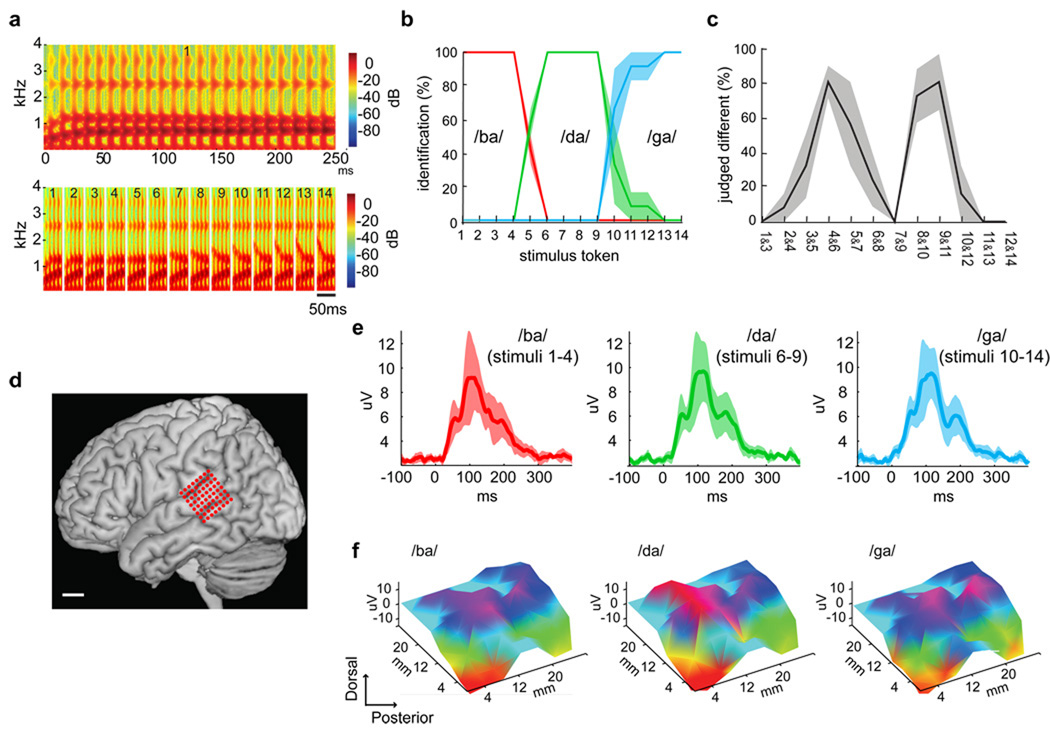
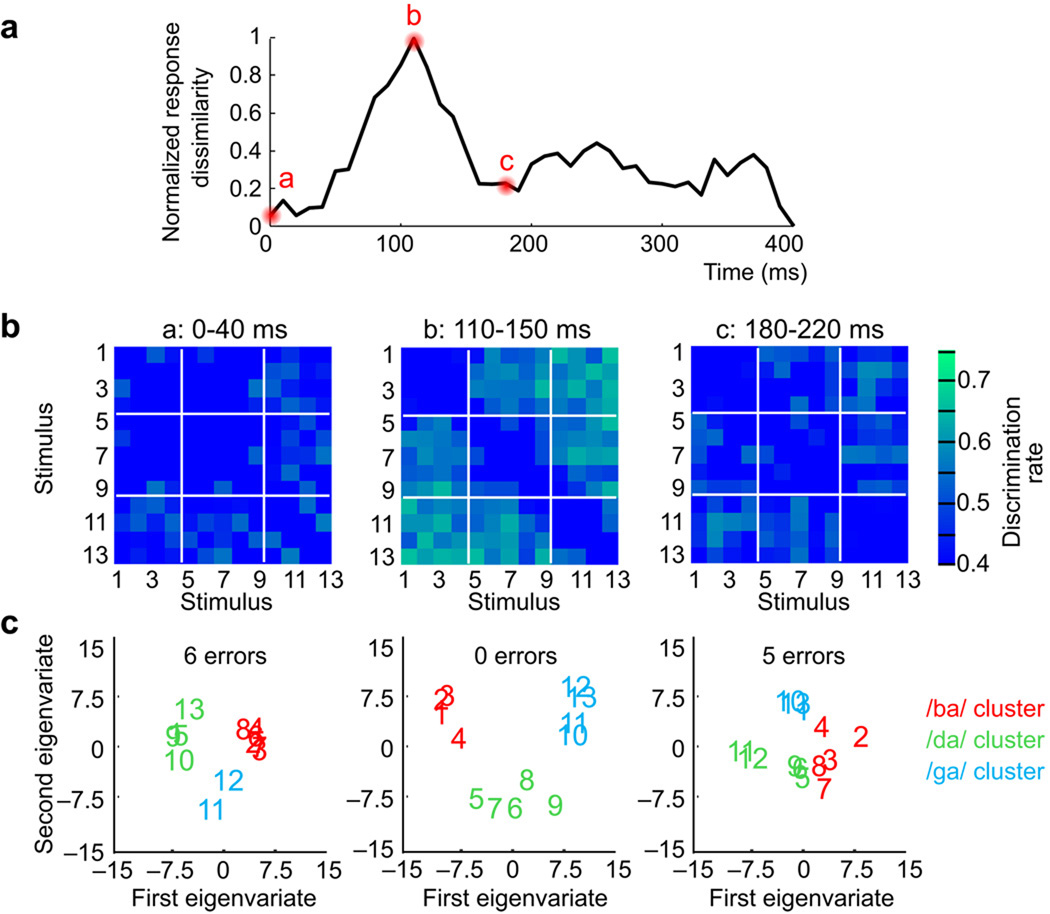
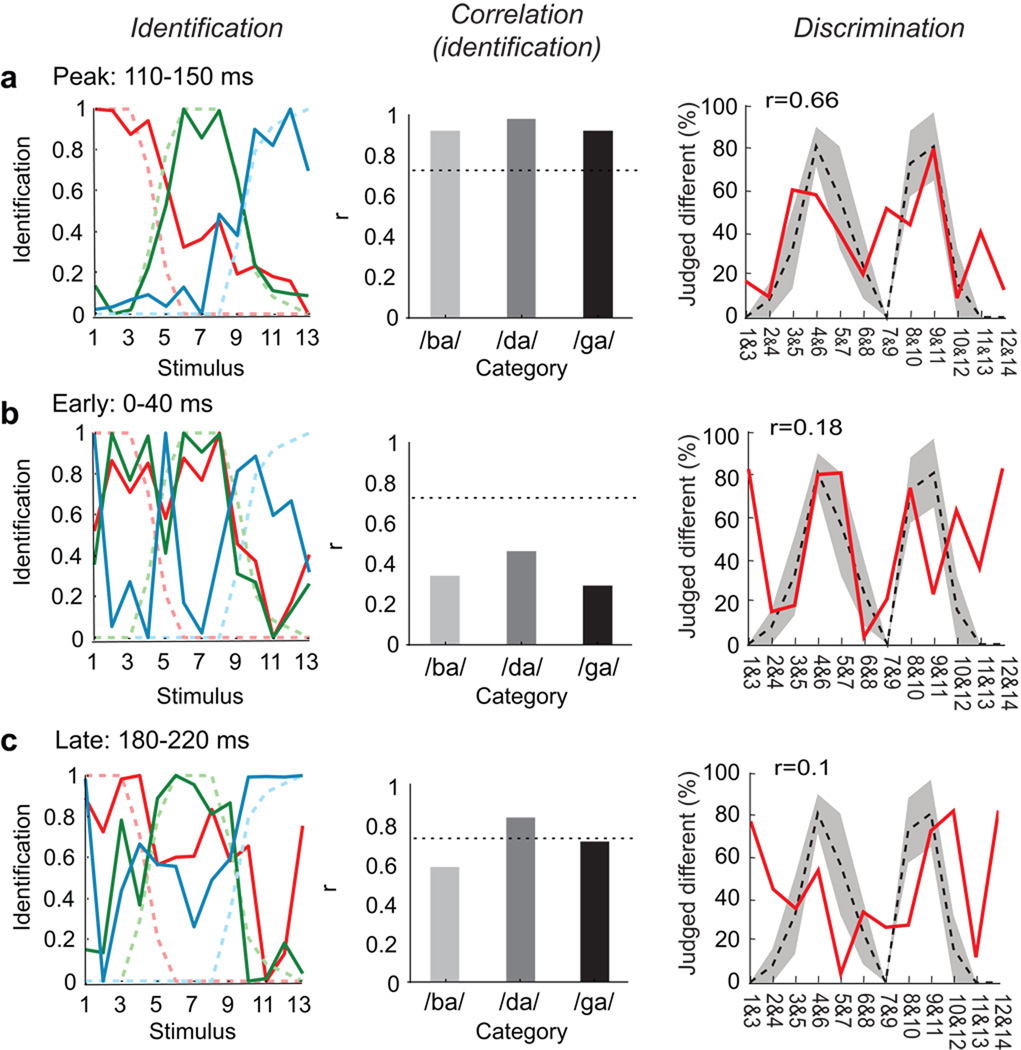
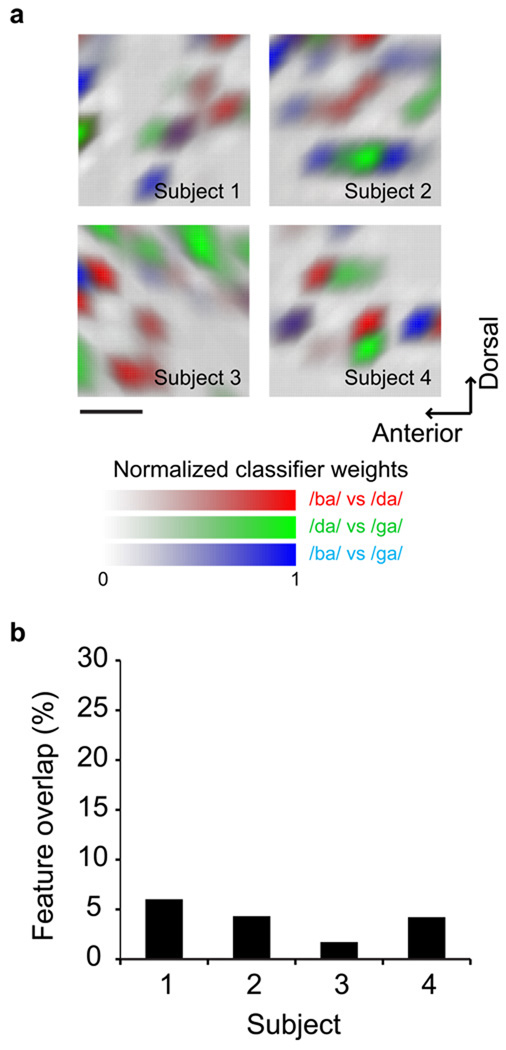
Speech perception requires the rapid and effortless extraction of meaningful phonetic information from a highly variable acoustic signal. A powerful example of this phenomenon is categorical speech perception, in which a continuum of acoustically varying sounds is transformed into perceptually distinct phoneme categories. We found that the neural representation of speech sounds is categorically organized in the human posterior superior temporal gyrus. Using intracranial high-density cortical surface arrays, we found that listening to synthesized speech stimuli varying in small and acoustically equal steps evoked distinct and invariant cortical population response patterns that were organized by their sensitivities to critical acoustic features. Phonetic category boundaries were similar between neurometric and psychometric functions. Although speech-sound responses were distributed, spatially discrete cortical loci were found to underlie specific phonetic discrimination. Our results provide direct evidence for acoustic-to-higher order phonetic level encoding of speech sounds in human language receptive cortex.
Figures
Comment in
-
Categorizing speech.Nat Neurosci. 2010 Nov;13(11):1304-6. doi: 10.1038/nn1110-1304. Nat Neurosci. 2010. PMID: 20975749 No abstract available.
References
-
- Perkell J, Klatt DH, editors. Invariance and variability in speech processes. Hillsdale, NJ: Lawrence Erlbaum Associates; 1986.
-
- Liberman AM, Cooper FS, Shankweiler DP, Studdert-Kennedy M. Perception of the speech code. Psychol Rev. 1967;74:431–461. - PubMed
-
- Diehl RL, Lotto AJ, Holt LL. Speech perception. Annu Rev Psychol. 2004;55:149–179. - PubMed
-
- Liberman AM, Mattingly IG. A specialization for speech perception. Science. 1989;243:489–494. - PubMed
-
- Vihman M. Phonological Development: The Origins of Language in the Child. Cambridge: Wiley-Blackwell; 1996.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
