

High resolution measurement of the glycolytic rate
- PMID: 20890447
- PMCID: PMC2947927
- DOI: 10.3389/fnene.2010.00026
High resolution measurement of the glycolytic rate
Abstract
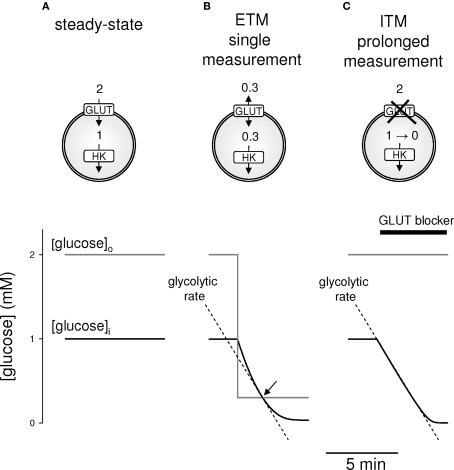
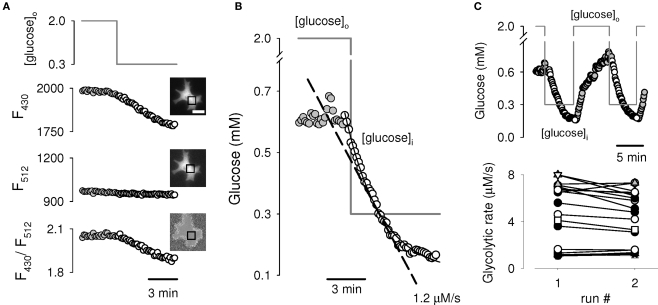
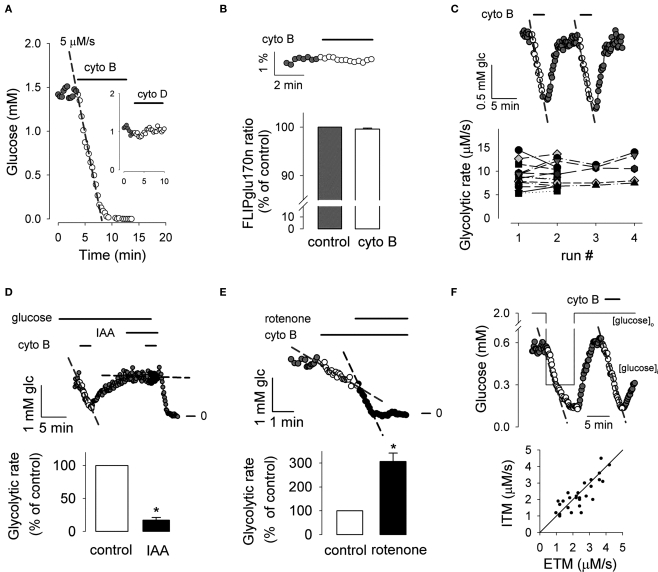
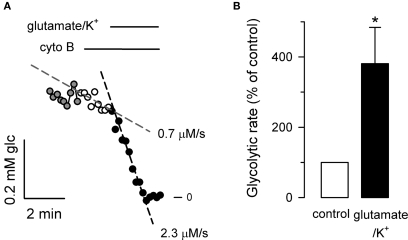
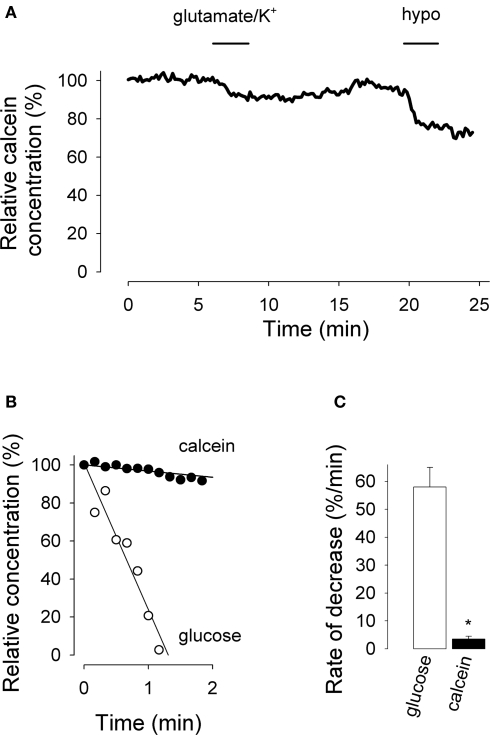
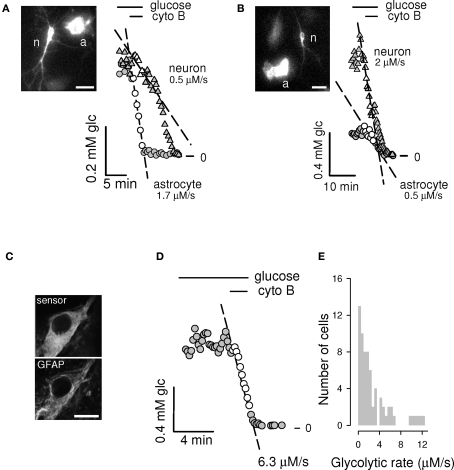
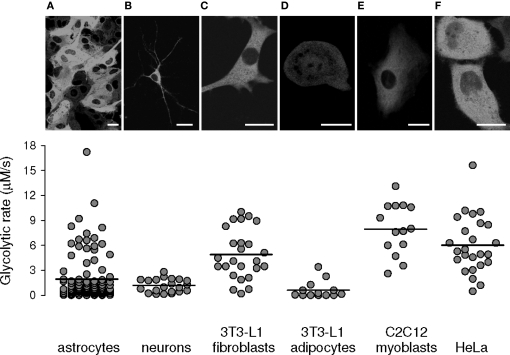
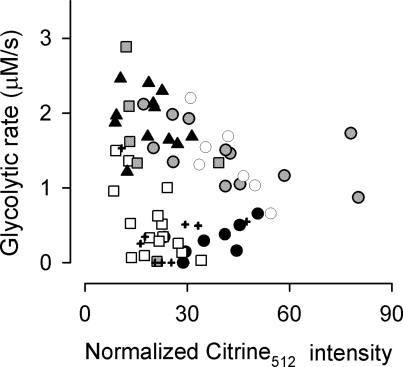
The glycolytic rate is sensitive to physiological activity, hormones, stress, aging, and malignant transformation. Standard techniques to measure the glycolytic rate are based on radioactive isotopes, are not able to resolve single cells and have poor temporal resolution, limitations that hamper the study of energy metabolism in the brain and other organs. A new method is described in this article, which makes use of a recently developed FRET glucose nanosensor to measure the rate of glycolysis in single cells with high temporal resolution. Used in cultured astrocytes, the method showed for the first time that glycolysis can be activated within seconds by a combination of glutamate and K(+), supporting a role for astrocytes in neurometabolic and neurovascular coupling in the brain. It was also possible to make a direct comparison of metabolism in neurons and astrocytes lying in close proximity, paving the way to a high-resolution characterization of brain energy metabolism. Single-cell glycolytic rates were also measured in fibroblasts, adipocytes, myoblasts, and tumor cells, showing higher rates for undifferentiated cells and significant metabolic heterogeneity within cell types. This method should facilitate the investigation of tissue metabolism at the single-cell level and is readily adaptable for high-throughput analysis.
Keywords: FRET nanosensor; cytochalasin B; glucose; glycolysis.
Figures
References
-
- Barros L. F., Bittner C. X., Loaiza A., Porras O. H. (2007). A quantitative overview of glucose dynamics in the gliovascular unit. Glia 55, 1222–1237 - PubMed
-
- Barros L. F., Courjaret R., Jakoby P., Loaiza A., Lohr C., Deitmer J. W. (2009). Preferential transport and metabolism of glucose in Bergmann glia over Purkinje cells: a multiphoton study of cerebellar slices. Glia 57, 962–970 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources