

STDP in Recurrent Neuronal Networks
- PMID: 20890448
- PMCID: PMC2947928
- DOI: 10.3389/fncom.2010.00023
STDP in Recurrent Neuronal Networks
Abstract
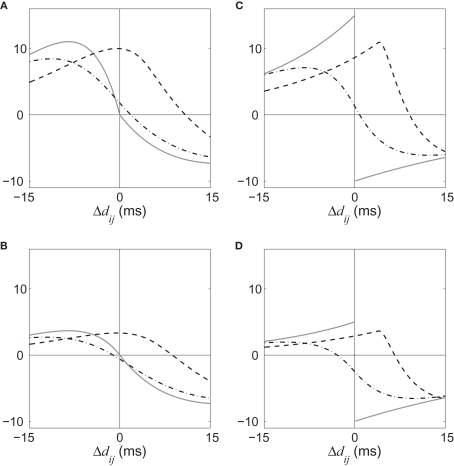
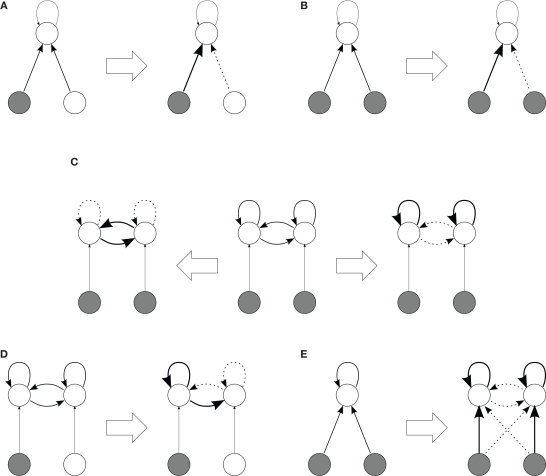
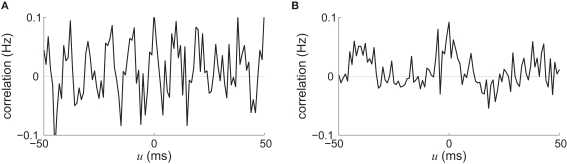
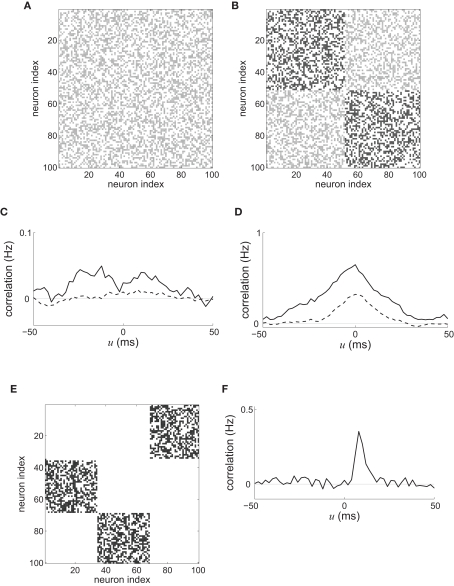
Recent results about spike-timing-dependent plasticity (STDP) in recurrently connected neurons are reviewed, with a focus on the relationship between the weight dynamics and the emergence of network structure. In particular, the evolution of synaptic weights in the two cases of incoming connections for a single neuron and recurrent connections are compared and contrasted. A theoretical framework is used that is based upon Poisson neurons with a temporally inhomogeneous firing rate and the asymptotic distribution of weights generated by the learning dynamics. Different network configurations examined in recent studies are discussed and an overview of the current understanding of STDP in recurrently connected neuronal networks is presented.
Keywords: STDP; network structure; recurrent neuronal network; self-organization / unsupervised learning; spike-time correlations.
Figures
Similar articles
-
Emergence of network structure due to spike-timing-dependent plasticity in recurrent neuronal networks. II. Input selectivity--symmetry breaking.Biol Cybern. 2009 Aug;101(2):103-14. doi: 10.1007/s00422-009-0320-y. Epub 2009 Jun 18. Biol Cybern. 2009. PMID: 19536559
-
Emergence of network structure due to spike-timing-dependent plasticity in recurrent neuronal networks. I. Input selectivity--strengthening correlated input pathways.Biol Cybern. 2009 Aug;101(2):81-102. doi: 10.1007/s00422-009-0319-4. Epub 2009 Jun 18. Biol Cybern. 2009. PMID: 19536560
-
Emergence of network structure due to spike-timing-dependent plasticity in recurrent neuronal networks IV: structuring synaptic pathways among recurrent connections.Biol Cybern. 2009 Dec;101(5-6):427-44. doi: 10.1007/s00422-009-0346-1. Epub 2009 Nov 24. Biol Cybern. 2009. PMID: 19937070
-
Propagation delays determine neuronal activity and synaptic connectivity patterns emerging in plastic neuronal networks.Chaos. 2018 Oct;28(10):106308. doi: 10.1063/1.5037309. Chaos. 2018. PMID: 30384625 Review.
-
Dendritic mechanisms controlling spike-timing-dependent synaptic plasticity.Trends Neurosci. 2007 Sep;30(9):456-63. doi: 10.1016/j.tins.2007.06.010. Epub 2007 Aug 31. Trends Neurosci. 2007. PMID: 17765330 Review.
Cited by
-
Emergence of connectivity motifs in networks of model neurons with short- and long-term plastic synapses.PLoS One. 2014 Jan 15;9(1):e84626. doi: 10.1371/journal.pone.0084626. eCollection 2014. PLoS One. 2014. PMID: 24454735 Free PMC article.
-
Effects of Firing Variability on Network Structures with Spike-Timing-Dependent Plasticity.Front Comput Neurosci. 2018 Jan 23;12:1. doi: 10.3389/fncom.2018.00001. eCollection 2018. Front Comput Neurosci. 2018. PMID: 29410621 Free PMC article.
-
Delay selection by spike-timing-dependent plasticity in recurrent networks of spiking neurons receiving oscillatory inputs.PLoS Comput Biol. 2013;9(2):e1002897. doi: 10.1371/journal.pcbi.1002897. Epub 2013 Feb 7. PLoS Comput Biol. 2013. PMID: 23408878 Free PMC article.
-
Spike-Timing-Dependent Plasticity Mediated by Dopamine and its Role in Parkinson's Disease Pathophysiology.Front Netw Physiol. 2022 Mar 4;2:817524. doi: 10.3389/fnetp.2022.817524. eCollection 2022. Front Netw Physiol. 2022. PMID: 36926058 Free PMC article. Review.
-
Decoupling of interacting neuronal populations by time-shifted stimulation through spike-timing-dependent plasticity.PLoS Comput Biol. 2023 Feb 1;19(2):e1010853. doi: 10.1371/journal.pcbi.1010853. eCollection 2023 Feb. PLoS Comput Biol. 2023. PMID: 36724144 Free PMC article.
References
-
- Amit D. J., Brunel N. (1997). Dynamics of a recurrent network of spiking neurons before and following learning. Netw. Comput. Neural Syst. 8, 373–40410.1088/0954-898X/8/4/003 - DOI
LinkOut - more resources
Full Text Sources