

Otoferlin is a calcium sensor that directly regulates SNARE-mediated membrane fusion
- PMID: 20921140
- PMCID: PMC2953437
- DOI: 10.1083/jcb.201002089
Otoferlin is a calcium sensor that directly regulates SNARE-mediated membrane fusion
Abstract
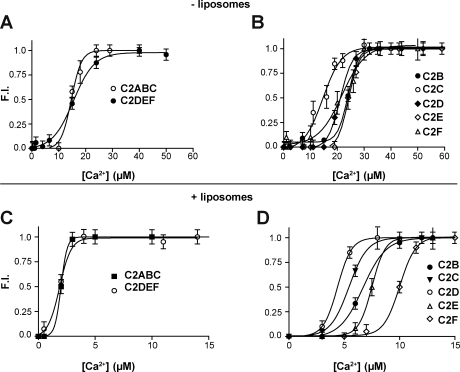
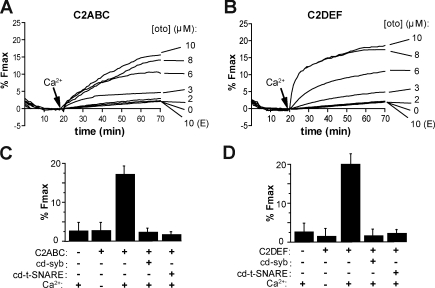
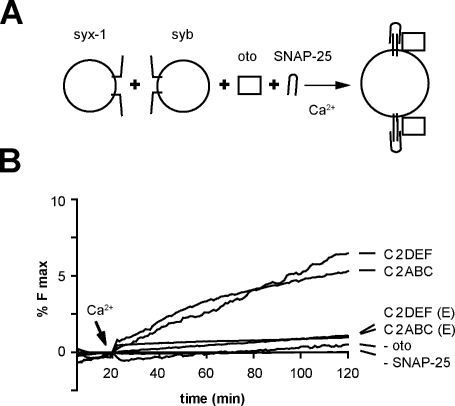
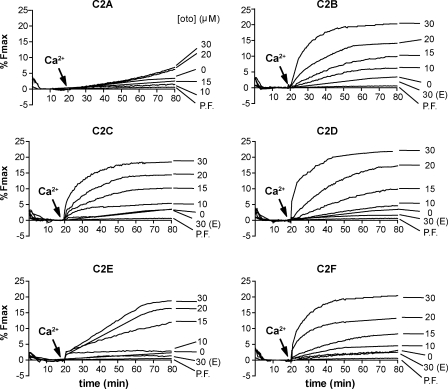
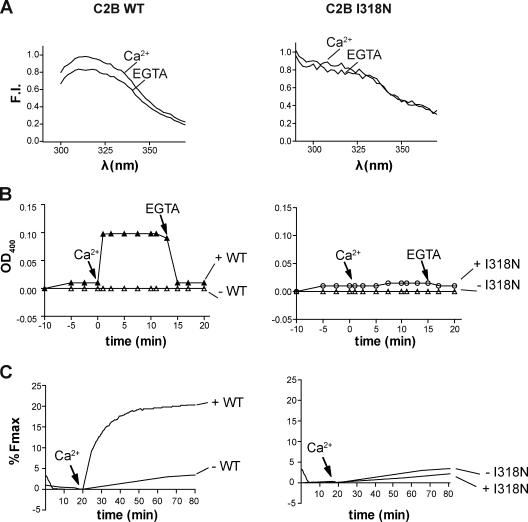
Otoferlin is a large multi-C2 domain protein proposed to act as a calcium sensor that regulates synaptic vesicle exocytosis in cochlear hair cells. Although mutations in otoferlin have been associated with deafness, its contribution to neurotransmitter release is unresolved. Using recombinant proteins, we demonstrate that five of the six C2 domains of otoferlin sense calcium with apparent dissociation constants that ranged from 13-25 µM; in the presence of membranes, these apparent affinities increase by up to sevenfold. Using a reconstituted membrane fusion assay, we found that five of the six C2 domains of otoferlin stimulate membrane fusion in a calcium-dependent manner. We also demonstrate that a calcium binding-deficient form of the C2C domain is incapable of stimulating membrane fusion, further underscoring the importance of calcium for the protein's function. These results demonstrate for the first time that otoferlin is a calcium sensor that can directly regulate soluble N-ethyl-maleimide sensitive fusion protein attachment protein receptor-mediated membrane fusion reactions.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
