

Protein turnover of the Wallenda/DLK kinase regulates a retrograde response to axonal injury
- PMID: 20921142
- PMCID: PMC2953441
- DOI: 10.1083/jcb.201006039
Protein turnover of the Wallenda/DLK kinase regulates a retrograde response to axonal injury
Abstract
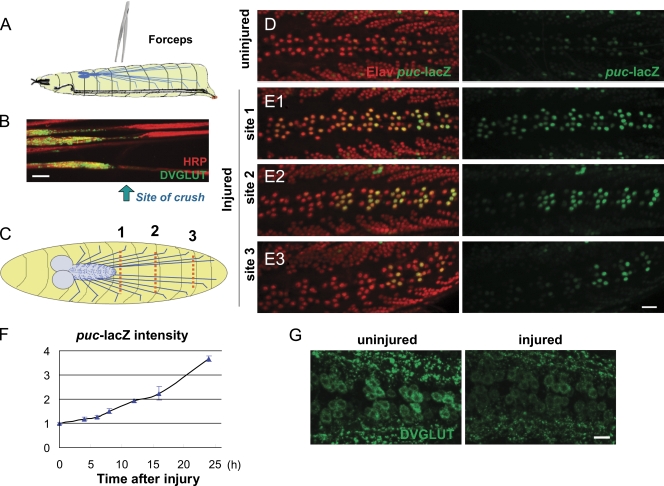
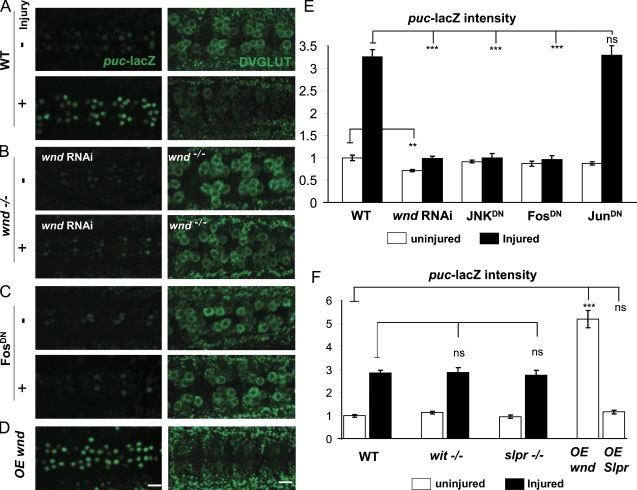
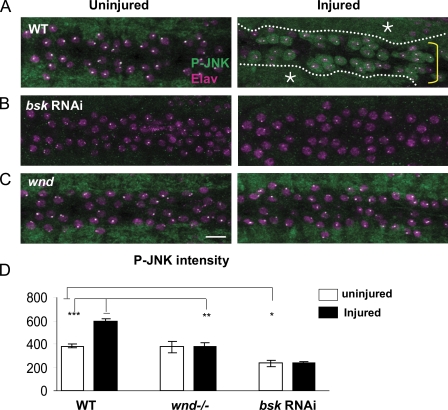
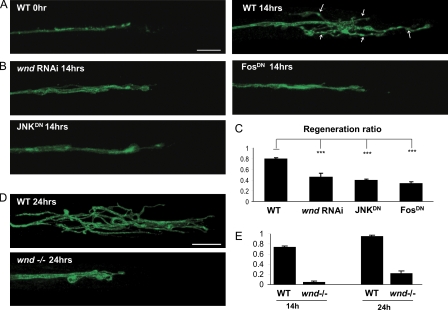
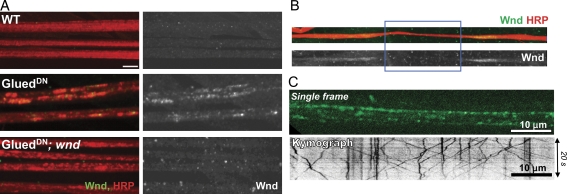
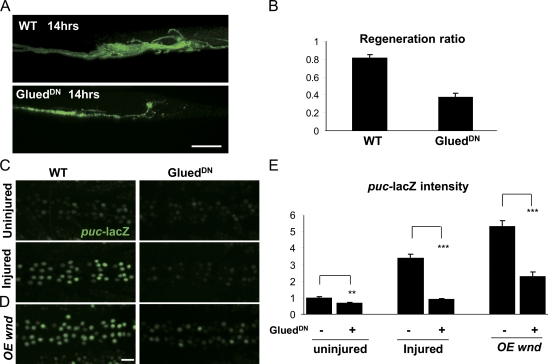
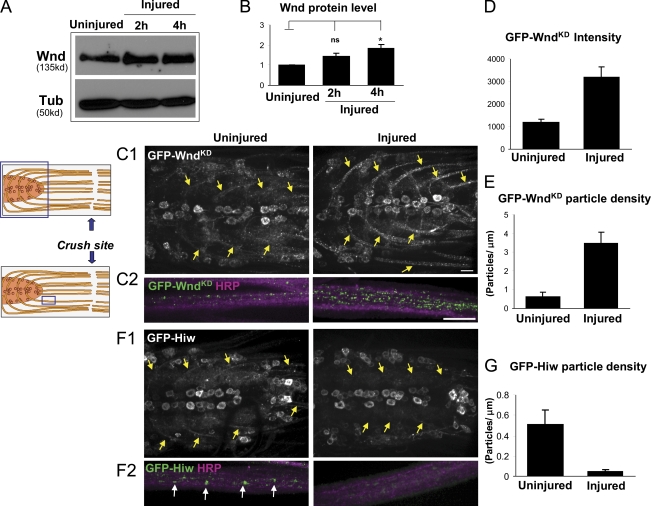
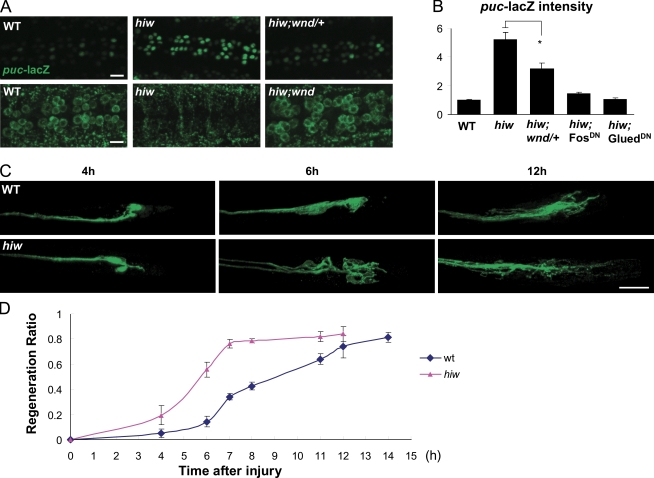
Regenerative responses to axonal injury involve changes in gene expression; however, little is known about how such changes can be induced from a distant site of injury. In this study, we describe a nerve crush assay in Drosophila melanogaster to study injury signaling and regeneration mechanisms. We find that Wallenda (Wnd), a conserved mitogen-activated protein kinase (MAPK) kinase kinase homologous to dual leucine zipper kinase, functions as an upstream mediator of a cell-autonomous injury signaling cascade that involves the c-Jun NH(2)-terminal kinase MAPK and Fos transcription factor. Wnd is physically transported in axons, and axonal transport is required for the injury signaling mechanism. Wnd is regulated by a conserved E3 ubiquitin ligase, named Highwire (Hiw) in Drosophila. Injury induces a rapid increase in Wnd protein concomitantly with a decrease in Hiw protein. In hiw mutants, injury signaling is constitutively active, and neurons initiate a faster regenerative response. Our data suggest that the regulation of Wnd protein turnover by Hiw can function as a damage surveillance mechanism for responding to axonal injury.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
