

Interaction of calcium-dependent activator protein for secretion 1 (CAPS1) with the class II ADP-ribosylation factor small GTPases is required for dense-core vesicle trafficking in the trans-Golgi network
- PMID: 20921225
- PMCID: PMC2992304
- DOI: 10.1074/jbc.M110.137414
Interaction of calcium-dependent activator protein for secretion 1 (CAPS1) with the class II ADP-ribosylation factor small GTPases is required for dense-core vesicle trafficking in the trans-Golgi network
Abstract
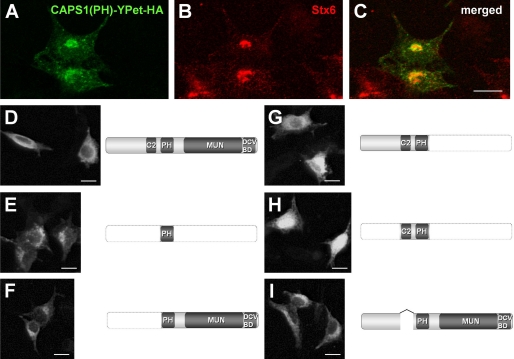
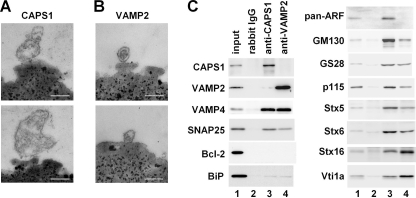
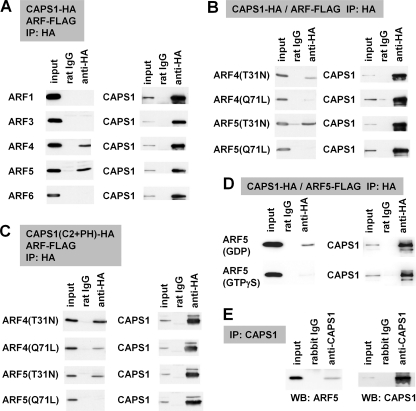
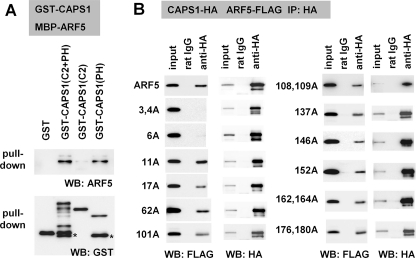
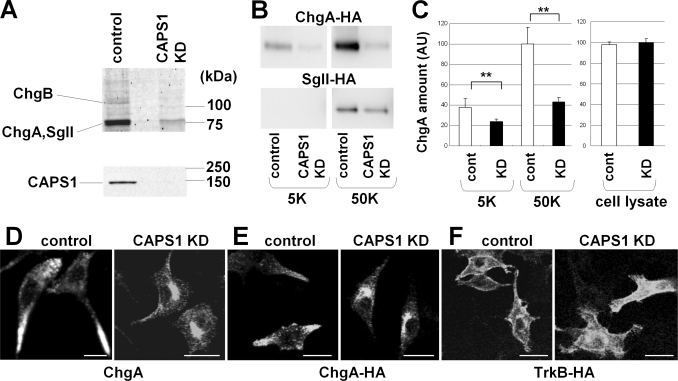
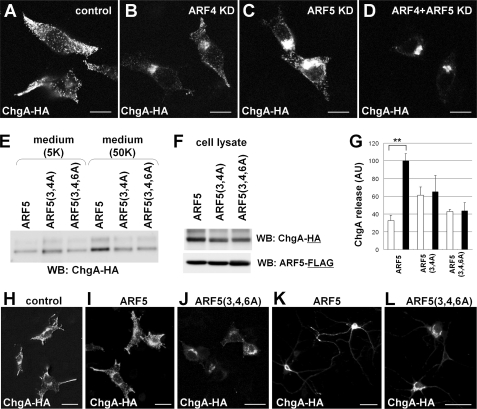
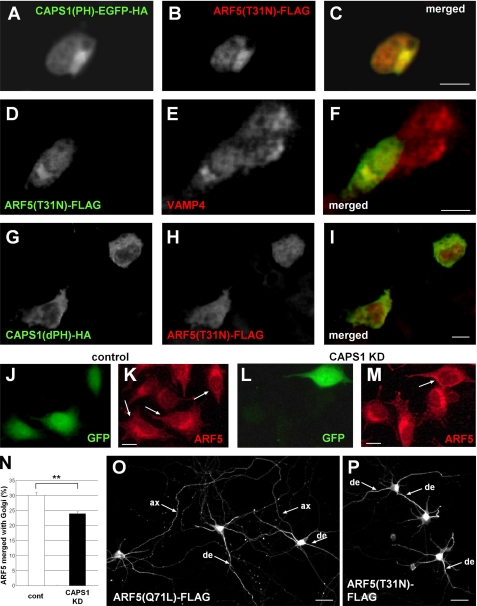
Ca(2+)-dependent activator protein for secretion (CAPS) regulates exocytosis of catecholamine- or neuropeptide-containing dense-core vesicles (DCVs) at secretion sites, such as nerve terminals. However, large amounts of CAPS protein are localized in the cell soma, and the role of somal CAPS protein remains unclear. The present study shows that somal CAPS1 plays an important role in DCV trafficking in the trans-Golgi network. The anti-CAPS1 antibody appeared to pull down membrane fractions, including many Golgi-associated proteins, such as ADP-ribosylation factor (ARF) small GTPases. Biochemical analyses of the protein-protein interaction showed that CAPS1 interacted specifically with the class II ARF4/ARF5, but not with other classes of ARFs, via the pleckstrin homology domain in a GDP-bound ARF form-specific manner. The pleckstrin homology domain of CAPS1 showed high affinity for the Golgi membrane, thereby recruiting ARF4/ARF5 to the Golgi complex. Knockdown of either CAPS1 or ARF4/ARF5 expression caused accumulation of chromogranin, a DCV marker protein, in the Golgi, thereby reducing its DCV secretion. In addition, the overexpression of CAPS1 binding-deficient ARF5 mutants induced aberrant chromogranin accumulation in the Golgi and consequently reduced its DCV secretion. These findings implicate a functional role for CAPS1 protein in the soma, a major subcellular localization site of CAPS1 in many cell types, in regulating DCV trafficking in the trans-Golgi network; this activity occurs via protein-protein interaction with ARF4/ARF5 in a GDP-dependent manner.
Figures
Similar articles
-
Calcium-dependent activator protein for secretion 2 interacts with the class II ARF small GTPases and regulates dense-core vesicle trafficking.FEBS J. 2012 Feb;279(3):384-94. doi: 10.1111/j.1742-4658.2011.08431.x. Epub 2011 Dec 12. FEBS J. 2012. PMID: 22111578
-
The priming factor CAPS1 regulates dense-core vesicle acidification by interacting with rabconnectin3β/WDR7 in neuroendocrine cells.J Biol Chem. 2019 Jun 14;294(24):9402-9415. doi: 10.1074/jbc.RA119.007504. Epub 2019 Apr 19. J Biol Chem. 2019. PMID: 31004036 Free PMC article.
-
CAPS1 deficiency perturbs dense-core vesicle trafficking and Golgi structure and reduces presynaptic release probability in the mouse brain.J Neurosci. 2013 Oct 30;33(44):17326-34. doi: 10.1523/JNEUROSCI.2777-13.2013. J Neurosci. 2013. PMID: 24174665 Free PMC article.
-
ARF proteins: roles in membrane traffic and beyond.Nat Rev Mol Cell Biol. 2006 May;7(5):347-58. doi: 10.1038/nrm1910. Nat Rev Mol Cell Biol. 2006. PMID: 16633337 Review.
-
Activation of toxin ADP-ribosyltransferases by eukaryotic ADP-ribosylation factors.Mol Cell Biochem. 1999 Mar;193(1-2):153-7. Mol Cell Biochem. 1999. PMID: 10331652 Review.
Cited by
-
Maternal immunoglobulin G affects brain development of mouse offspring.J Neuroinflammation. 2024 May 2;21(1):114. doi: 10.1186/s12974-024-03100-z. J Neuroinflammation. 2024. PMID: 38698428 Free PMC article.
-
CAPS-1 promotes fusion competence of stationary dense-core vesicles in presynaptic terminals of mammalian neurons.Elife. 2015 Feb 26;4:e05438. doi: 10.7554/eLife.05438. Elife. 2015. PMID: 25719439 Free PMC article.
-
Morpho-functional architecture of the Golgi complex of neuroendocrine cells.Front Endocrinol (Lausanne). 2013 Mar 28;4:41. doi: 10.3389/fendo.2013.00041. eCollection 2013. Front Endocrinol (Lausanne). 2013. PMID: 23543640 Free PMC article.
-
Kinetics of Arf1 inactivation regulates Golgi organisation and function in non-adherent fibroblasts.Biol Open. 2023 Apr 15;12(4):bio059669. doi: 10.1242/bio.059669. Epub 2023 May 4. Biol Open. 2023. PMID: 36946871 Free PMC article.
-
LAMP5 in presynaptic inhibitory terminals in the hindbrain and spinal cord: a role in startle response and auditory processing.Mol Brain. 2019 Mar 12;12(1):20. doi: 10.1186/s13041-019-0437-4. Mol Brain. 2019. PMID: 30867010 Free PMC article.
References
-
- Renden R., Berwin B., Davis W., Ann K., Chin C. T., Kreber R., Ganetzky B., Martin T. F., Broadie K. (2001) Neuron 31, 421–437 - PubMed
-
- Berwin B., Floor E., Martin T. F. (1998) Neuron 21, 137–145 - PubMed
-
- Tandon A., Bannykh S., Kowalchyk J. A., Banerjee A., Martin T. F., Balch W. E. (1998) Neuron 21, 147–154 - PubMed
-
- Speidel D., Varoqueaux F., Enk C., Nojiri M., Grishanin R. N., Martin T. F., Hofmann K., Brose N., Reim K. (2003) J. Biol. Chem. 278, 52802–52809 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
