

Optimization of enzymatic reaction conditions for generating representative pools of cDNA from small RNA
- PMID: 20921270
- PMCID: PMC2995414
- DOI: 10.1261/rna.2242610
Optimization of enzymatic reaction conditions for generating representative pools of cDNA from small RNA
Abstract
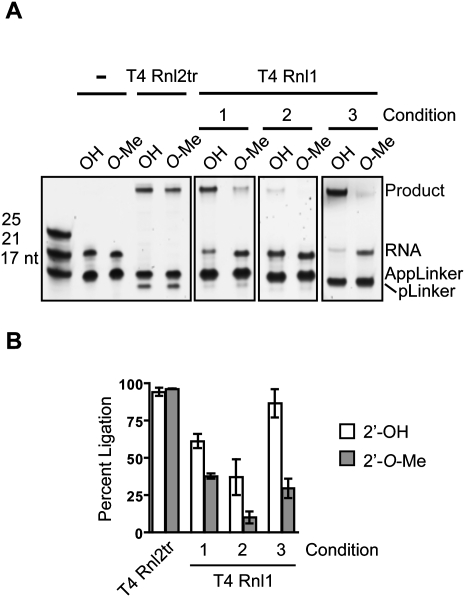
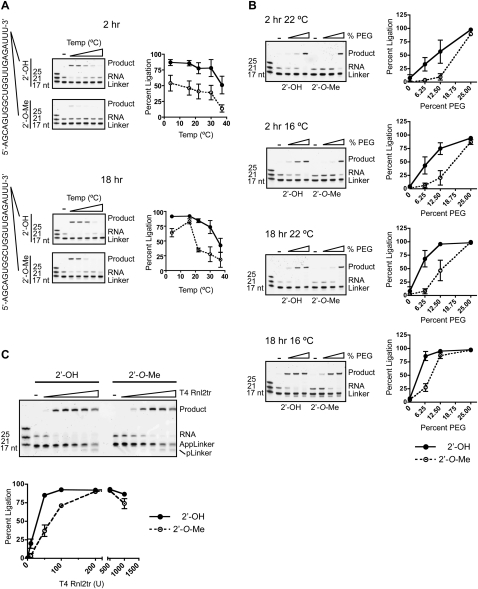
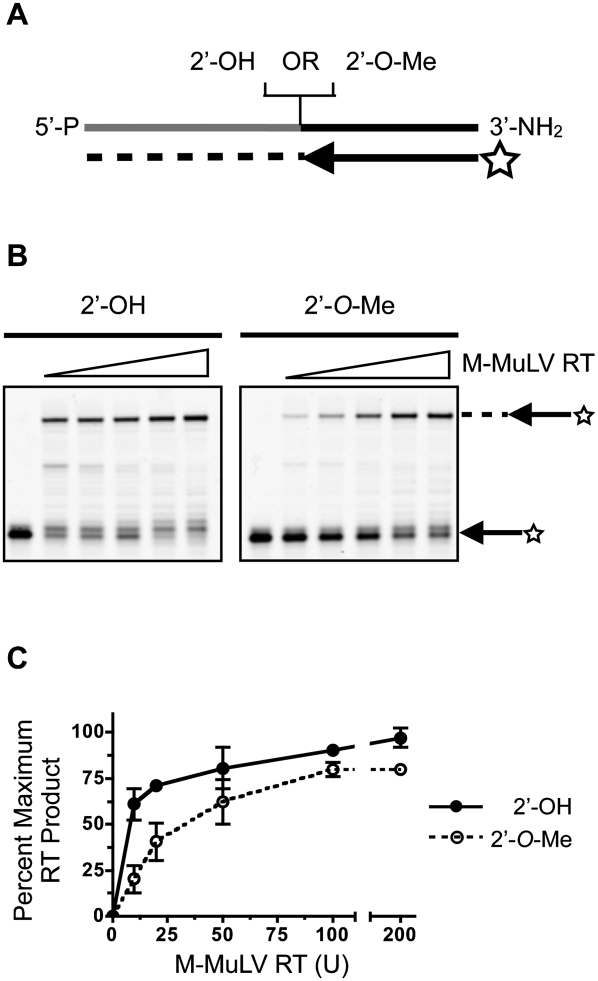
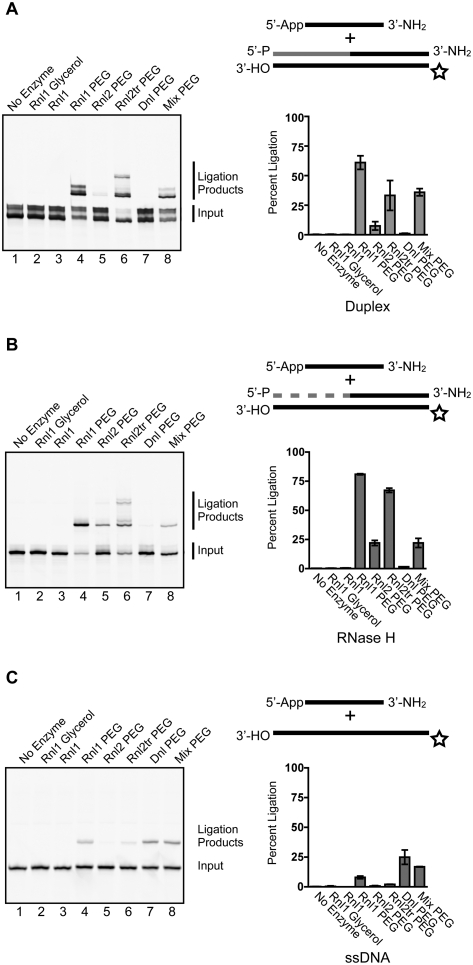
Small regulatory RNA repertoires in biological samples are heterogeneous mixtures that may include species arising from varied biosynthetic pathways and modification events. Small RNA profiling and discovery approaches ought to capture molecules in a way that is representative of expression level. It follows that the effects of RNA modifications on representation should be minimized. The collection of high-quality, representative data, therefore, will be highly dependent on bias-free sample manipulation in advance of quantification. We examined the impact of 2'-O-methylation of the 3'-terminal nucleotide of small RNA on key enzymatic reactions of standard front-end manipulation schemes. Here we report that this common modification negatively influences the representation of these small RNA species. Deficits occurred at multiple steps as determined by gel analysis of synthetic input RNA and by quantification and sequencing of derived cDNA pools. We describe methods to minimize the effects of 2'-O-methyl modification of small RNA 3'-termini using T4 RNA ligase 2 truncated, and other optimized reaction conditions, demonstrating their use by quantifying representation of miRNAs and piRNAs in cDNA pools prepared from biological samples.
Figures



References
-
- Aravin A, Tuschl T 2005. Identification and characterization of small RNAs involved in RNA silencing. FEBS Lett 579: 5830–5840 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources