

Endocytosis is required for Toll signaling and shaping of the Dorsal/NF-kappaB morphogen gradient during Drosophila embryogenesis
- PMID: 20921412
- PMCID: PMC2964194
- DOI: 10.1073/pnas.1009157107
Endocytosis is required for Toll signaling and shaping of the Dorsal/NF-kappaB morphogen gradient during Drosophila embryogenesis
Abstract
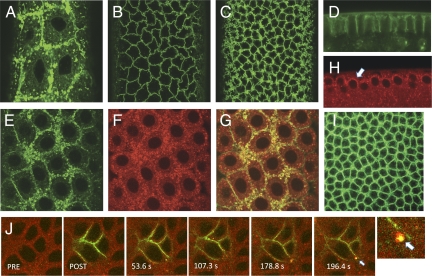
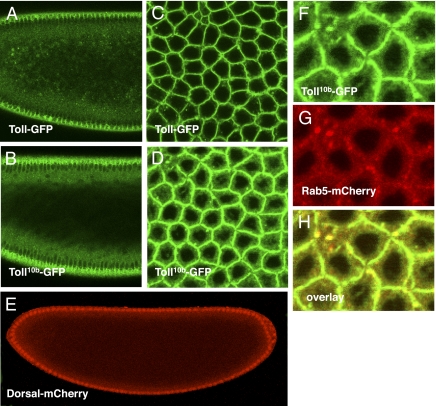
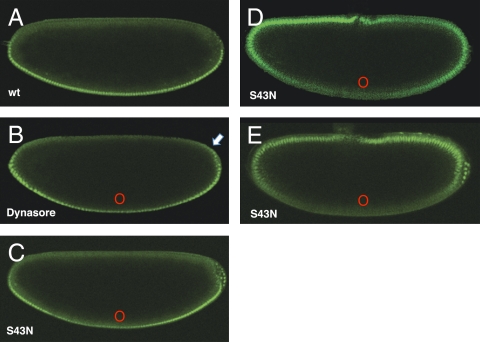
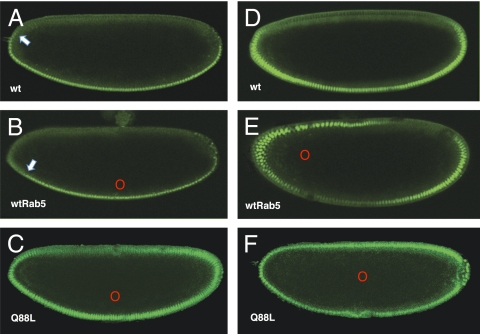
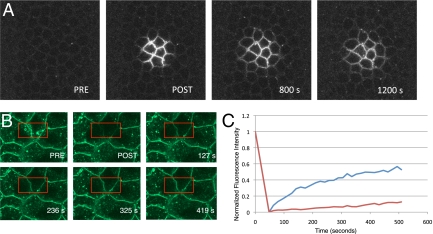
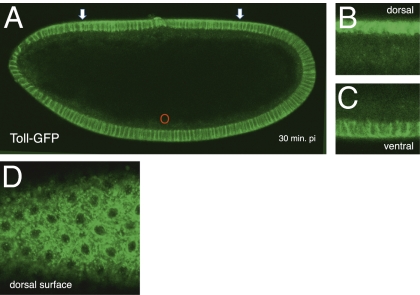
Dorsoventral cell fate in the Drosophila embryo is specified by activation of the Toll receptor, leading to a ventral-to-dorsal gradient across nuclei of the NF-κB transcription factor Dorsal. Toll receptor has been investigated genetically, molecularly, and immunohistologically, but much less is known about its dynamics in living embryos. Using live imaging of fluorescent protein chimeras, we find that Toll is recruited from the plasma membrane to Rab5(+) early endosomes. The distribution of a constitutively active form of Toll, Toll(10b), is shifted from the plasma membrane to early endosomes. Inhibition of endocytosis on the ventral side of the embryo attenuates Toll signaling ventrally and causes Dorsal to accumulate on the dorsal side of the embryo, essentially inverting the dorsal/ventral axis. Conversely, enhancing endocytosis laterally greatly potentiates Toll signaling locally, altering the shape of the Dorsal gradient. Photoactivation and fluorescence recovery after photobleaching studies reveal that Toll exhibits extremely limited lateral diffusion within the plasma membrane, whereas Toll is highly compartmentalized in endosomes. When endocytosis is blocked ventrally, creating an ectopic dorsal signaling center, Toll is preferentially endocytosed at the ectopic signaling center. We propose that Toll signals from an endocytic compartment rather than the plasma membrane. Our studies reveal that endocytosis plays a pivotal role in the spatial regulation of Toll receptor activation and signaling and in the correct shaping of the nuclear Dorsal concentration gradient.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Shedding light on Toll signaling through live imaging.Fly (Austin). 2011 Apr-Jun;5(2):141-6. doi: 10.4161/fly.5.2.14768. Epub 2011 Apr 1. Fly (Austin). 2011. PMID: 21273815
-
Dorsoventral axis formation in the Drosophila embryo--shaping and transducing a morphogen gradient.Curr Biol. 2005 Nov 8;15(21):R887-99. doi: 10.1016/j.cub.2005.10.026. Curr Biol. 2005. PMID: 16271864 Review.
-
Robustness of the Dorsal morphogen gradient with respect to morphogen dosage.PLoS Comput Biol. 2020 Apr 6;16(4):e1007750. doi: 10.1371/journal.pcbi.1007750. eCollection 2020 Apr. PLoS Comput Biol. 2020. PMID: 32251432 Free PMC article.
-
Vertebrate Rel proteins exhibit Dorsal-like activities in early Drosophila embryogenesis.Dev Dyn. 2006 Apr;235(4):949-57. doi: 10.1002/dvdy.20713. Dev Dyn. 2006. PMID: 16493693
-
Formation, interpretation, and regulation of the Drosophila Dorsal/NF-κB gradient.Curr Top Dev Biol. 2020;137:143-191. doi: 10.1016/bs.ctdb.2019.11.007. Epub 2019 Dec 26. Curr Top Dev Biol. 2020. PMID: 32143742 Review.
Cited by
-
A Toll-receptor map underlies structural brain plasticity.Elife. 2020 Feb 18;9:e52743. doi: 10.7554/eLife.52743. Elife. 2020. PMID: 32066523 Free PMC article.
-
Polydnaviral ankyrin proteins aid parasitic wasp survival by coordinate and selective inhibition of hematopoietic and immune NF-kappa B signaling in insect hosts.PLoS Pathog. 2013;9(8):e1003580. doi: 10.1371/journal.ppat.1003580. Epub 2013 Aug 29. PLoS Pathog. 2013. PMID: 24009508 Free PMC article.
-
PmAP2-β depletion enhanced activation of the Toll signaling pathway during yellow head virus infection in the black tiger shrimp Penaeus monodon.Sci Rep. 2021 May 18;11(1):10534. doi: 10.1038/s41598-021-89922-w. Sci Rep. 2021. PMID: 34006863 Free PMC article.
-
Drosophila melanogaster Toll-9 elicits antiviral immunity against Drosophila C virus.J Virol. 2025 Jun 17;99(6):e0221424. doi: 10.1128/jvi.02214-24. Epub 2025 May 14. J Virol. 2025. PMID: 40366172 Free PMC article.
-
Genetic Screen in Drosophila Larvae Links ird1 Function to Toll Signaling in the Fat Body and Hemocyte Motility.PLoS One. 2016 Jul 28;11(7):e0159473. doi: 10.1371/journal.pone.0159473. eCollection 2016. PLoS One. 2016. PMID: 27467079 Free PMC article.
References
-
- DeLotto R, Spierer P. A gene required for the specification of dorsal-ventral pattern in Drosophila appears to encode a serine protease. Nature. 1986;323:688–692. - PubMed
-
- DeLotto Y, DeLotto R. Proteolytic processing of the Drosophila Spätzle protein by easter generates a dimeric NGF-like molecule with ventralising activity. Mech Dev. 1998;72:141–148. - PubMed
-
- Moussian B, Roth S. Dorsoventral axis formation in the Drosophila embryo—Shaping and transducing a morphogen gradient. Curr Biol. 2005;15(Suppl):R887–R899. - PubMed
-
- Charatsi I, Luschnig S, Bartoszewski S, Nüsslein-Volhard C, Moussian B. Krapfen/dMyd88 is required for the establishment of dorsoventral pattern in the Drosophila embryo. Mech Dev. 2003;120:219–226. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
