

Error-prone DNA repair activity during somatic hypermutation in shark B lymphocytes
- PMID: 20921520
- PMCID: PMC2959145
- DOI: 10.4049/jimmunol.1000779
Error-prone DNA repair activity during somatic hypermutation in shark B lymphocytes
Abstract
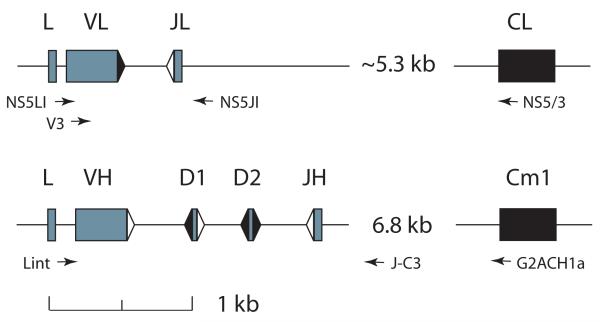
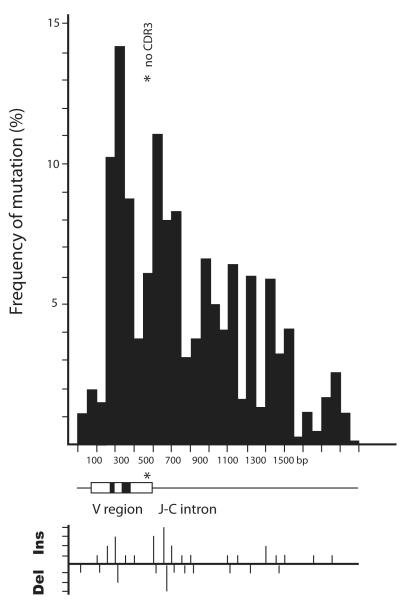
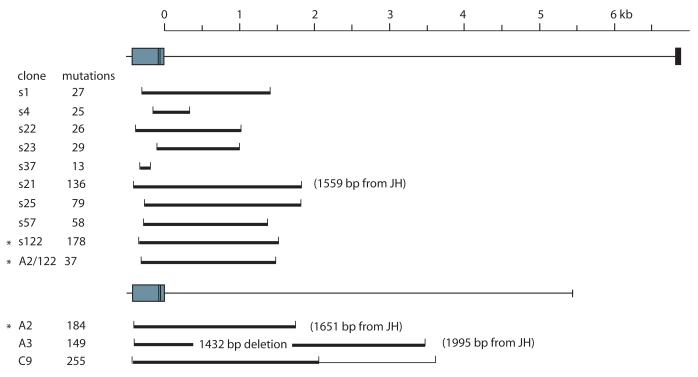
Sharks are representatives of the earliest vertebrates that possess an immune system utilizing V(D)J recombination to generate Ag receptors. Their Ab repertoire diversity is based in part on a somatic hypermutation process that introduces adjacent nucleotide substitutions of 2-5 bp. We have isolated mutant nonfunctional Ig rearrangements and intronic flank sequences to characterize the nonselected, intrinsic properties of this phenomenon; changes unique to shark were observed. Duplications and deletions were associated with N additions, suggesting participation of a DNA polymerase with some degree of template independence during the repair of DNA breaks initiated by activation-induced cytidine deaminase. Other mutations were consistent with some in vitro activities of mammalian translesion DNA polymerase η: tandem base substitutions, strand slippage, and small insertions/deletions. The nature of substitution patterns shows that DNA lesions at shark Ig genes recruit DNA repair factors with a species-specific repertoire of activities. We speculate that the tandem mutations are introduced by direct sequential misinsertions and that, in shark B cells, the mispairs tend to be extended rather than proofread. Despite extensive changes undergone by some mutants, the physical range of mutational activity remained restricted to VDJ and within the first 2-kb portion of the 6.8-kb J-C intron, perhaps a self-regulating aspect of activation-induced cytidine deaminase action that is conserved in evolution.
Figures



Similar articles
-
The multiple shark Ig H chain genes rearrange and hypermutate autonomously.J Immunol. 2011 Sep 1;187(5):2492-501. doi: 10.4049/jimmunol.1101671. Epub 2011 Jul 29. J Immunol. 2011. PMID: 21804022 Free PMC article.
-
Somatic hypermutation of TCR γ V genes in the sandbar shark.Dev Comp Immunol. 2012 May;37(1):176-83. doi: 10.1016/j.dci.2011.08.018. Epub 2011 Sep 8. Dev Comp Immunol. 2012. PMID: 21925537
-
DNA targets of AID evolutionary link between antibody somatic hypermutation and class switch recombination.Adv Immunol. 2009;101:163-89. doi: 10.1016/S0065-2776(08)01005-5. Adv Immunol. 2009. PMID: 19231595 Review.
-
The Complex Interplay between DNA Injury and Repair in Enzymatically Induced Mutagenesis and DNA Damage in B Lymphocytes.Int J Mol Sci. 2017 Aug 30;18(9):1876. doi: 10.3390/ijms18091876. Int J Mol Sci. 2017. PMID: 28867784 Free PMC article. Review.
-
The translesion DNA polymerase theta plays a dominant role in immunoglobulin gene somatic hypermutation.EMBO J. 2005 Nov 2;24(21):3757-69. doi: 10.1038/sj.emboj.7600833. Epub 2005 Oct 13. EMBO J. 2005. PMID: 16222339 Free PMC article.
Cited by
-
Ancient Use of Ig Variable Domains Contributes Significantly to the TCRδ Repertoire.J Immunol. 2019 Sep 1;203(5):1265-1275. doi: 10.4049/jimmunol.1900369. Epub 2019 Jul 24. J Immunol. 2019. PMID: 31341077 Free PMC article.
-
Origin of immunoglobulin isotype switching.Curr Biol. 2012 May 22;22(10):872-80. doi: 10.1016/j.cub.2012.03.060. Epub 2012 Apr 26. Curr Biol. 2012. PMID: 22542103 Free PMC article.
-
Heavy chain-only antibody genes in fish evolved to generate unique CDR3 repertoire.Eur J Immunol. 2022 Feb;52(2):247-260. doi: 10.1002/eji.202149588. Epub 2021 Nov 19. Eur J Immunol. 2022. PMID: 34708869 Free PMC article.
-
Lost structural and functional inter-relationships between Ig and TCR loci in mammals revealed in sharks.Immunogenetics. 2021 Feb;73(1):17-33. doi: 10.1007/s00251-020-01183-5. Epub 2021 Jan 15. Immunogenetics. 2021. PMID: 33449123 Free PMC article. Review.
-
The multiple shark Ig H chain genes rearrange and hypermutate autonomously.J Immunol. 2011 Sep 1;187(5):2492-501. doi: 10.4049/jimmunol.1101671. Epub 2011 Jul 29. J Immunol. 2011. PMID: 21804022 Free PMC article.
References
-
- Du Pasquier L, Wilson M, Greenberg AS, Flajnik MF. Somatic mutation in ectothermic vertebrates: musings on selection and origins. Curr. Top. Microbiol. Immunol. 1998;229:199–216. - PubMed
-
- Muramatsu M, Kinoshita K, Fagarasan S, Yamada S, Shinkai Y, Honjo T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 2000;102:553–563. - PubMed
-
- Arakawa H, Hauschild J, Buerstedde JM. Requirement of the activation-induced deaminase (AID) gene for immunoglobulin gene conversion. Science. 2002;295:1301–1306. - PubMed
-
- Harris RS, Sale JE, Petersen-Mahrt SK, Neuberger MS. AID is essential for immunoglobulin V gene conversion in a cultured B cell line. Curr. Biol. 2002;12:435–8. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
