

Localization of an accessory helicase at the replisome is critical in sustaining efficient genome duplication
- PMID: 20923786
- PMCID: PMC3035471
- DOI: 10.1093/nar/gkq889
Localization of an accessory helicase at the replisome is critical in sustaining efficient genome duplication
Abstract
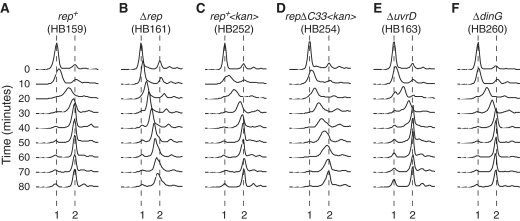
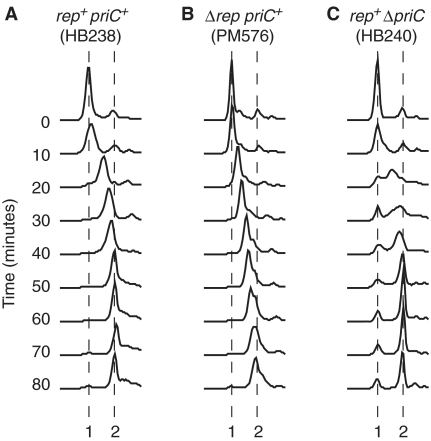
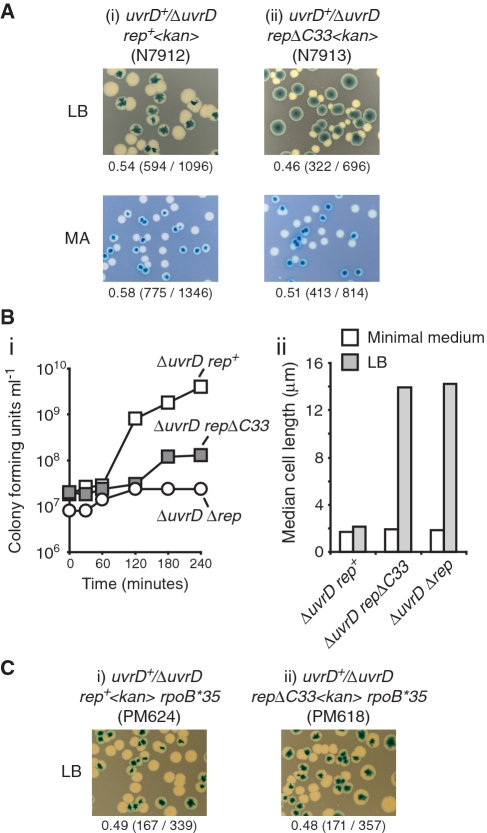
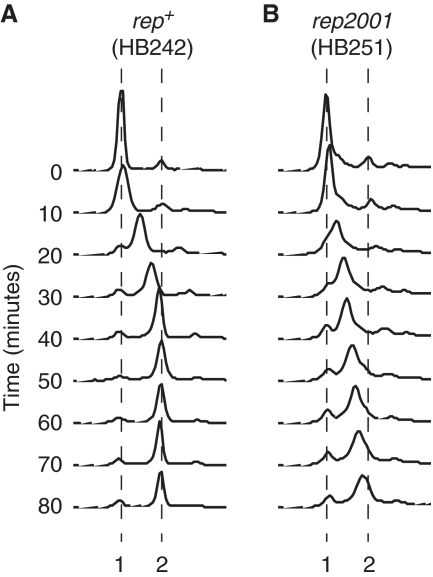
Genome duplication requires accessory helicases to displace proteins ahead of advancing replication forks. Escherichia coli contains three helicases, Rep, UvrD and DinG, that might promote replication of protein-bound DNA. One of these helicases, Rep, also interacts with the replicative helicase DnaB. We demonstrate that Rep is the only putative accessory helicase whose absence results in an increased chromosome duplication time. We show also that the interaction between Rep and DnaB is required for Rep to maintain rapid genome duplication. Furthermore, this Rep-DnaB interaction is critical in minimizing the need for both recombinational processing of blocked replication forks and replisome reassembly, indicating that colocalization of Rep and DnaB minimizes stalling and subsequent inactivation of replication forks. These data indicate that E. coli contains only one helicase that acts as an accessory motor at the fork in wild-type cells, that such an activity is critical for the maintenance of rapid genome duplication and that colocalization with the replisome is crucial for this function. Given that the only other characterized accessory motor, Saccharomyces cerevisiae Rrm3p, associates physically with the replisome, our demonstration of the functional importance of such an association indicates that colocalization may be a conserved feature of accessory replicative motors.
Figures

References
-
- French S. Consequences of replication fork movement through transcription units in vivo. Science. 1992;258:1362–1365. - PubMed
-
- Ivessa AS, Lenzmeier BA, Bessler JB, Goudsouzian LK, Schnakenberg SL, Zakian VA. The Saccharomyces cerevisiae helicase Rrm3p facilitates replication past nonhistone protein-DNA complexes. Mol. Cell. 2003;12:1525–1536. - PubMed
-
- Matson SW, Bean DW, George JW. DNA helicases: enzymes with essential roles in all aspects of DNA metabolism. Bioessays. 1994;16:13–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- BB/E020690/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/G005915/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0800970/MRC_/Medical Research Council/United Kingdom
- BB/E0020690/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
