

Maintenance of neuronal laterality in Caenorhabditis elegans through MYST histone acetyltransferase complex components LSY-12, LSY-13 and LIN-49
- PMID: 20923973
- PMCID: PMC2998328
- DOI: 10.1534/genetics.110.123661
Maintenance of neuronal laterality in Caenorhabditis elegans through MYST histone acetyltransferase complex components LSY-12, LSY-13 and LIN-49
Abstract
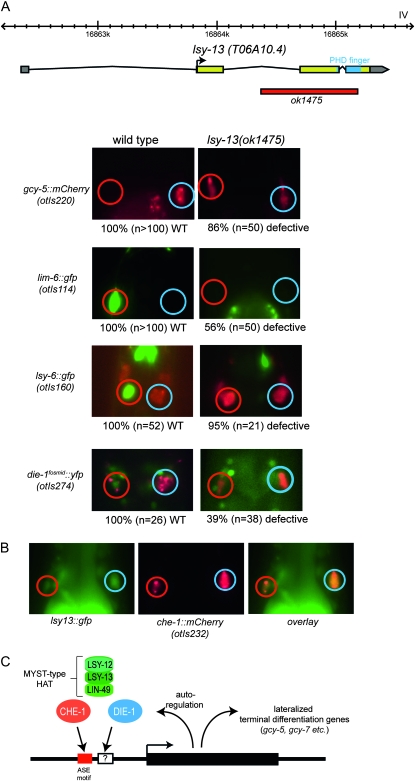
Left/right asymmetrically expressed genes permit an animal to perform distinct tasks with the right vs. left side of its brain. Once established during development, lateralized gene expression patterns need to be maintained during the life of the animal. We show here that a histone modifying complex, composed of the LSY-12 MYST-type histone acetyltransferase, the ING-family PHD domain protein LSY-13, and PHD/bromodomain protein LIN-49, is required to first initiate and then actively maintain lateralized gene expression in the gustatory system of the nematode Caenorhabditis elegans. Similar defects are observed upon postembryonic removal of two C2H2 zinc finger transcription factors, die-1 and che-1, demonstrating that a combination of transcription factors, which recognize DNA in a sequence-specific manner, and a histone modifying enzyme complex are responsible for inducing and maintaining neuronal laterality.
Figures


References
-
- Blau, H. M., 1992. Differentiation requires continuous active control. Annu. Rev. Biochem. 61 1213–1230. - PubMed
-
- Ceol, C. J., and H. R. Horvitz, 2004. A new class of C. elegans synMuv genes implicates a Tip60/NuA4-like HAT complex as a negative regulator of Ras signaling. Dev. Cell 6 563–576. - PubMed
-
- Chang, S., R. J. Johnston, C. Frokjaer-Jensen, S. Lockery and O. Hobert, 2004. MicroRNAs act sequentially and asymmetrically to control chemosensory laterality in the nematode. Nature 430 785–789. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
