

Review
doi: 10.1177/0022034510377791.
Epub 2010 Oct 5.
Aging and bone
Affiliations
- PMID: 20924069
- PMCID: PMC2991386
- DOI: 10.1177/0022034510377791
Item in Clipboard
Review
Aging and bone
J Dent Res.
2010 Dec.
Abstract
Bones provide mechanical and protective function, while also serving as housing for marrow and a site for regulation of calcium ion homeostasis. The properties of bones do not remain constant with age; rather, they change throughout life, in some cases improving in function, but in others, function deteriorates. Here we review the modifications in the mechanical function and shape of bones, the bone cells, the matrix they produce, and the mineral that is deposited on this matrix, while presenting recent theories about the factors leading to these changes.
Figures
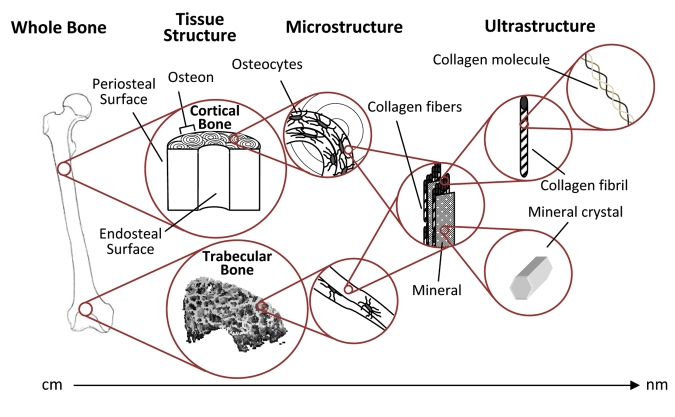
The structural levels of bone. Cortical bone is made up of longitudinally oriented osteons, and the trabecular bone within the metaphyses is made up of connected struts and plates. In both bone types, the bone is laid down in layers (lamellae). Both tissue types contain identical components, and their properties are dependent on the amount, morphology, and interaction of these components at each level.
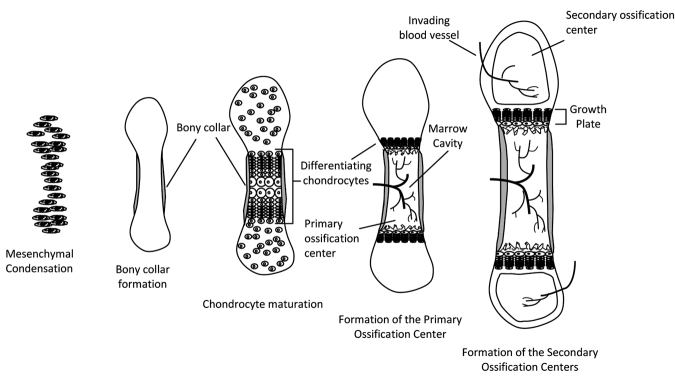
Endochondral ossification of long bones. Condensation of mesenchymal cells forms the general shape of the long bone. Differentiation of these cells into chondrocytes begins the process of bone formation in the primary ossification center. In late differentiation, chondrocytes undergo apoptosis, leaving behind a mineralized scaffold onto which osteoblasts brought in by the invading vasculature lay down bone, lengthening the bone while forming the marrow cavity. As development continues, secondary ossification centers form in the epiphyses. Bone increases in width through deposition of bone on the periosteal side and, through endochondral ossification, continues to increase in length until the growth plates fuse.
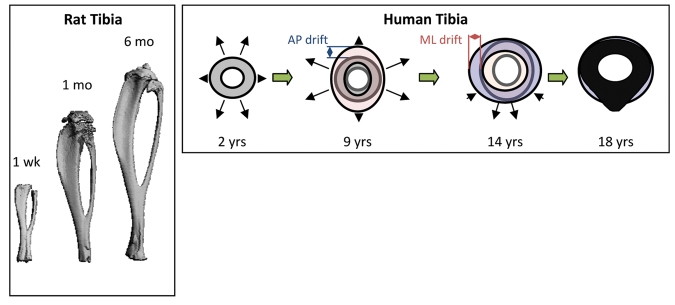
Long bone geometrical changes with age. Left: micro-CT 3D renderings of rat tibias with increasing age. Right: Cortical drift in the tibias of human bones with age. Changes in periosteal and endosteal remodeling around the bone are non-uniform throughout life. Note that the AP drift is larger from 2 to 9 yrs, and that the ML drift is greater from 9 to 14 yrs. Right panel adapted from Goldman et al. (2009). AP: anterior-posterior direction. ML: medial-lateral direction.
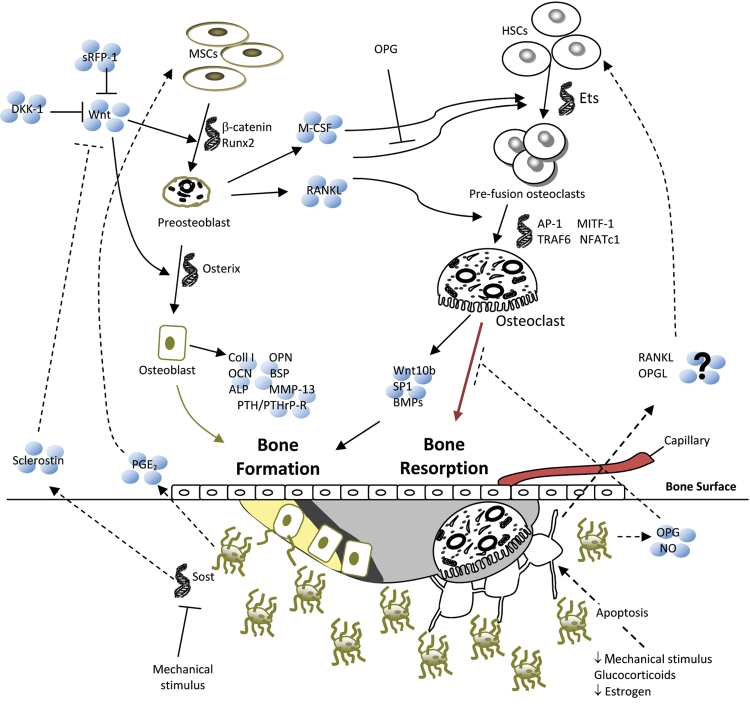
Bone remodeling pathways. Several factors can initiate bone remodeling, which, in a balanced system, begins with resorption by osteoclasts and ends with formation by osteoblasts. Signals from external sources (Wnts) and from within the bone (osteocyte apoptosis) contribute to osteoblast and osteoclast differentiation and activity, a selection of which is shown in this figure. Light gray = resorption pit, dark gray = osteoid, yellow = newly mineralized osteoid.
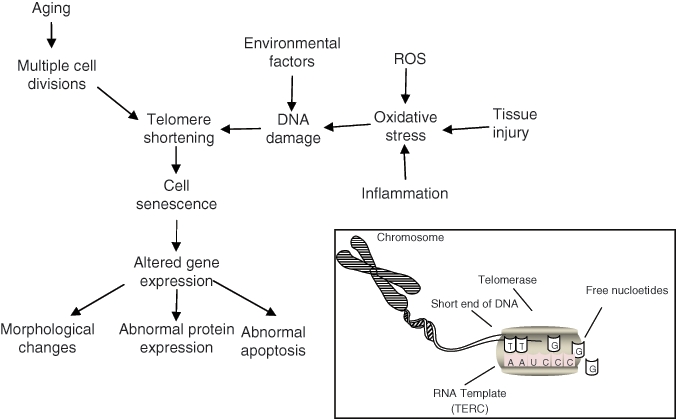
Factors affecting cell senescence. Cells reach senescence when regions at the ends of chromosomes, telomeres, reach a critical length due to the cells’ inability to replicate them properly. The inset shows the activity of an enzyme, telomerase, that allows stem cells to extend the length of the telomeres and therefore maintain an almost indefinite capacity for self-renewal. There are many factors that also contribute to telomere shortening, which may lead to abnormal cell behavior. The accumulation of damaged cells and the inability of a depleting stem cell population to replace these cells with aging may affect tissue function. Adapted from Muller (2009).
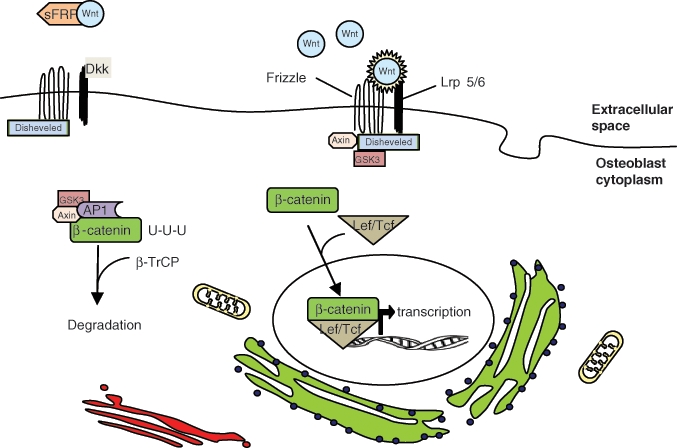
Wnt/β-catenin pathway of osteoblast differentiation. The translation of genes that stimulate osteoblast differentiation require the translocation of β-catenin from the cytoplasm to the nucleus of mesenchymal stem cells. β-catenin is targeted for degradation by the GSK3/AP1/Axin complex. Wnt binding to frizzled and Lrp 5/6 interrupts this process, allowing β-catenin to complex with Lef/Tcf. Dkk inhibits Wnt binding by interacting with Lrp 5/6. sFRP also inhibits this pathway by sequestering Wnt.
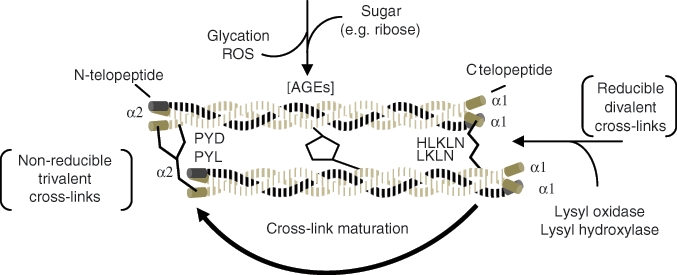
Chronological formation of collagen cross-links. As collagen matures, reducible cross-links become non-reducible. Advanced glycation end-products (AGEs) also accumulate between the helical parts of the molecules as the collagen persists in the tissue. Both contribute to the stiffening of the collagen matrix with age, which may contribute to bone tissue properties. Adapted from Leeming et al. (2009) and Saito and Marumo (2010).
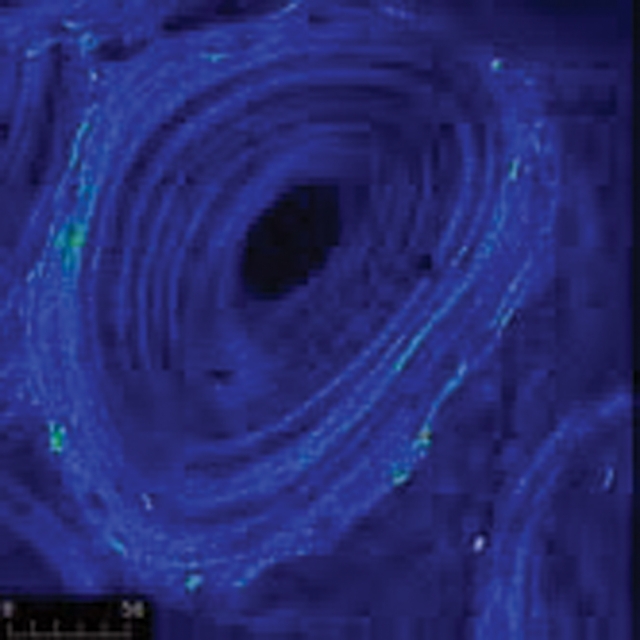
Collagen orientation of osteons obtained by second-harmonic generation microscopy, where the intensity is proportional to the orientation of the collagen. The orientation increases with tissue age, as is apparent in the outer, older rings of the osteon, which have a brighter intensity. Image from a 6-year-old baboon courtesy of Jayme Burket.
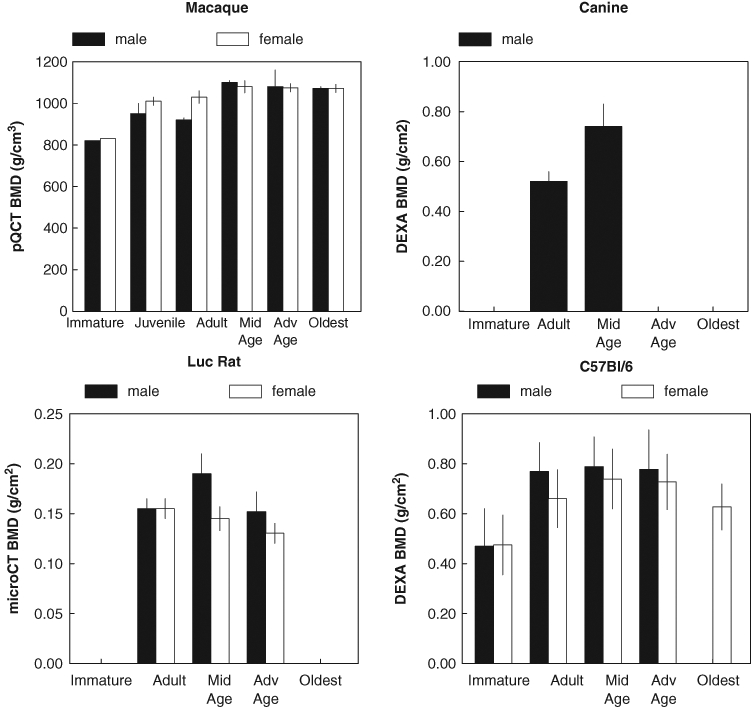
Change in BMD of various species with age. All of the plotted species show an increase in BMD with age. Male increases faster and female decreases earlier, but both sexes decline with age. Adapted from the following: macaque – Cerroni et al. (2000); canine – Schneider et al. (2004); Luc rat – Duque et al. (2009); C57Bl/6 – Somerville et al. (2004).
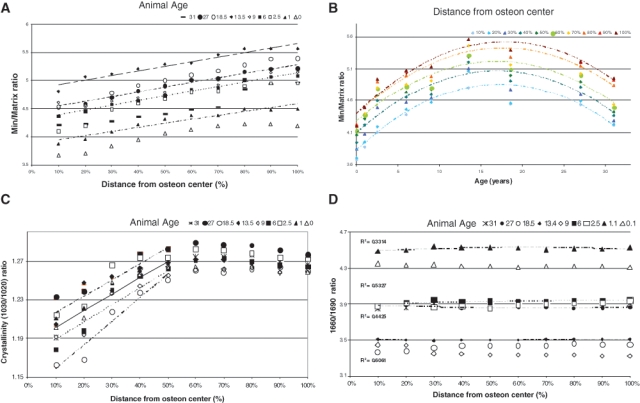
Heterogeneity of FTIR parameters in baboon osteonal bone. (A) Mineral/matrix ratio as a function of tissue age (distance from osteon center) in baboons of ages ranging from 0 to 37 yrs (R2 ~ 0.92-0.99). (B) Mineral/matrix ratio plotted as a function of baboon age for each section along the line from the center of the osteon (R2 ~ 0.83-0.89). (C) Crystallinity for each baboon sample as a function of tissue age (distance from the osteon center) (0.84 < R2 < 0.94). (D) Collagen cross-link ratio (maturity) for each baboon sample (0.1 < R2 < 0.5) as a function of tissue age (distance from the osteon center). Modified from Gourion-Arsiquaud et al. (2009b).
References
-
- Aaron JE, Makins NB, Sagreiya K. (1987). The microanatomy of trabecular bone loss in normal aging men and women. Clin Orthop Relat Res 215:260-271 - PubMed
-
- Adachi T, Aonuma Y, Ito S, Tanaka M, Hojo M, Takano-Yamamoto T, et al. (2009). Osteocyte calcium signaling response to bone matrix deformation. J Biomech 42:2507-2512 - PubMed
-
- Akkus O, Polyakova-Akkus A, Adar F, Schaffler MB. (2003). Aging of microstructural compartments in human compact bone. J Bone Miner Res 18:1012-1019 - PubMed
-
- Almeida M, Han L, Martin-Millan M, O’Brien CA, Manolagas SC. (2007b). Oxidative stress antagonizes Wnt signaling in osteoblast precursors by diverting beta-catenin from T cell factor- to forkhead box O-mediated transcription. J Biol Chem 282:27298-27305 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
