

Control of exocytosis by synaptotagmins and otoferlin in auditory hair cells
- PMID: 20926654
- PMCID: PMC3088501
- DOI: 10.1523/JNEUROSCI.2528-10.2010
Control of exocytosis by synaptotagmins and otoferlin in auditory hair cells
Abstract
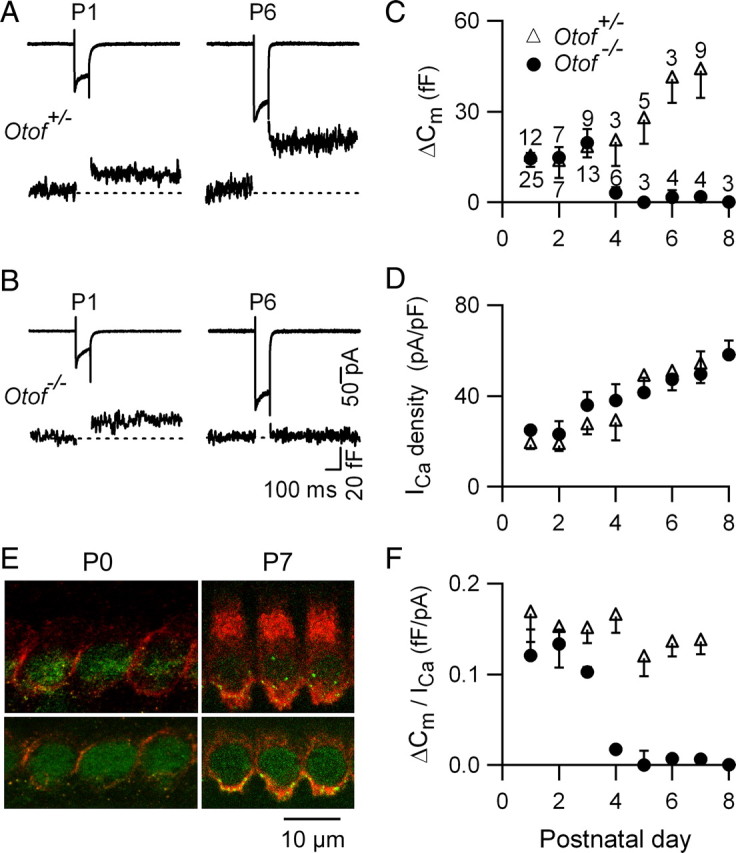
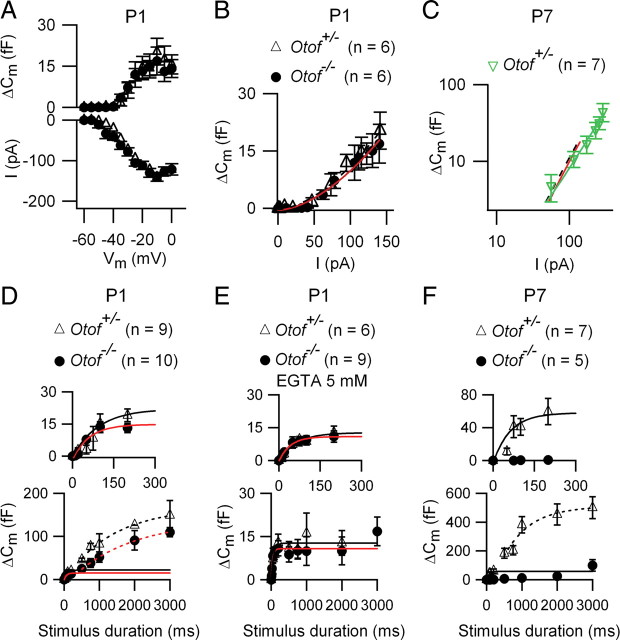
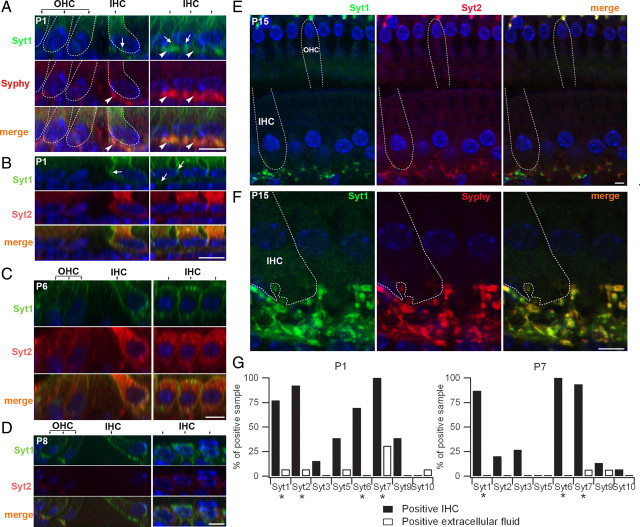
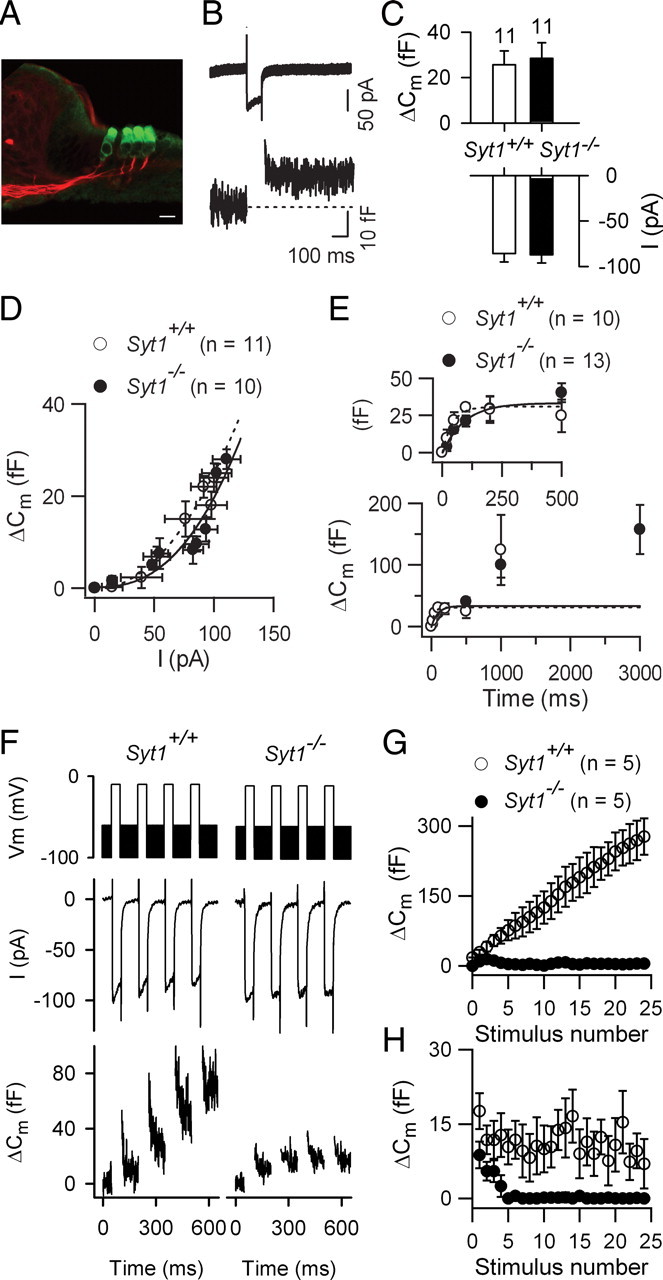
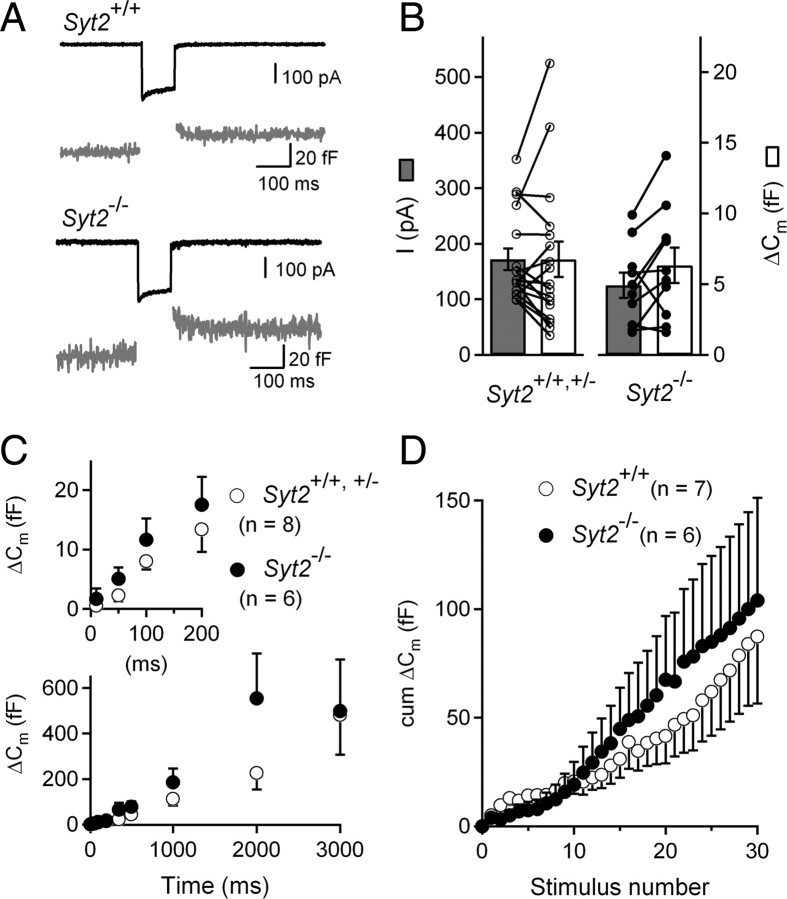
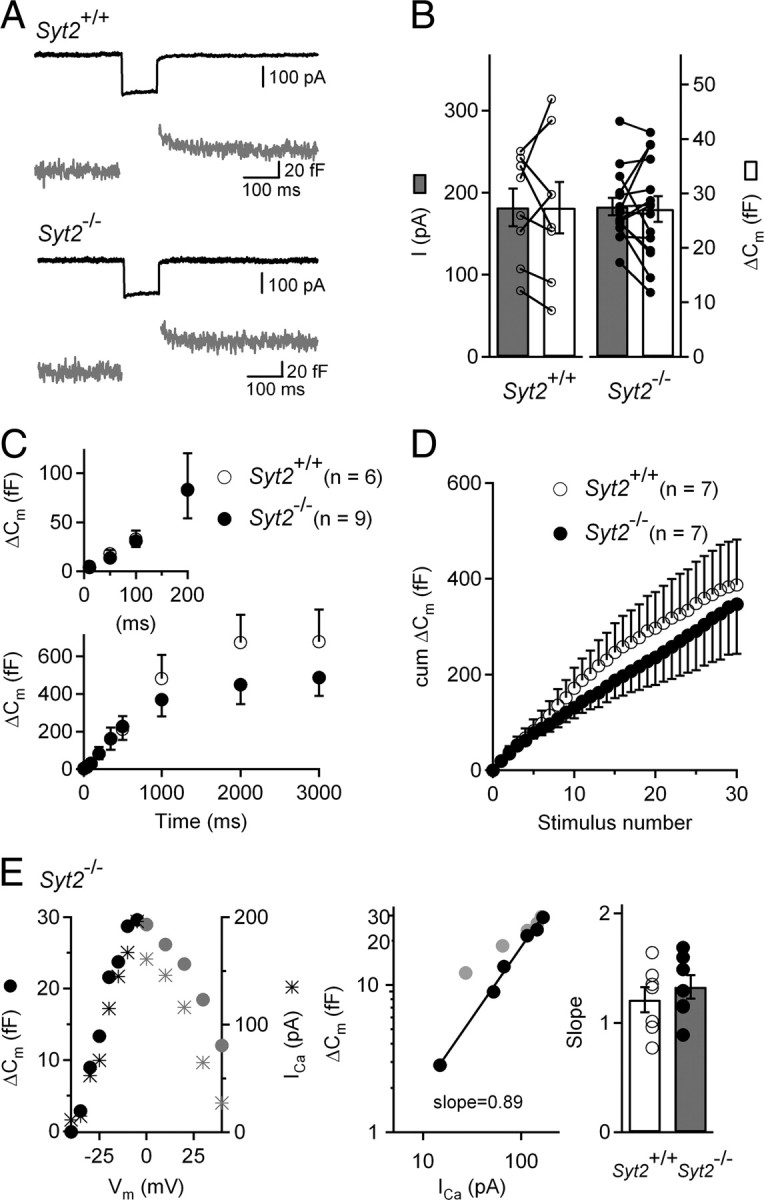
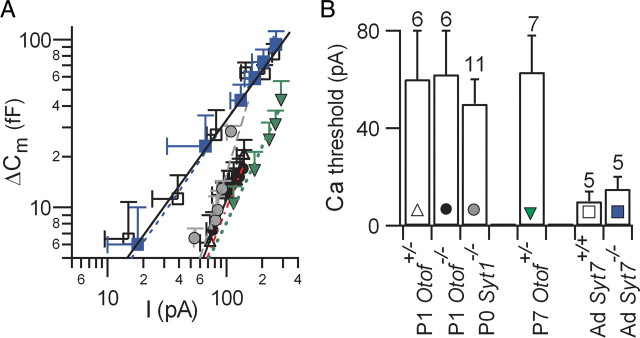
In pre-hearing mice, vesicle exocytosis at cochlear inner hair cell (IHC) ribbon synapses is triggered by spontaneous Ca(2+) spikes. At the onset of hearing, IHC exocytosis is then exclusively driven by graded potentials, and is characterized by higher Ca(2+) efficiency and improved synchronization of vesicular release. The molecular players involved in this transition are still unknown. Here we addressed the involvement of synaptotagmins and otoferlin as putative Ca(2+) sensors in IHC exocytosis during postnatal maturation of the cochlea. Using cell capacitance measurements, we showed that Ca(2+)-evoked exocytosis in mouse IHCs switches from an otoferlin-independent to an otoferlin-dependent mechanism at postnatal day 4. During this early exocytotic period, several synaptotagmins (Syts), including Syt1, Syt2 and Syt7, were detected in IHCs. The exocytotic response as well as the release of the readily releasable vesicle pool (RRP) was, however, unchanged in newborn mutant mice lacking Syt1, Syt2 or Syt7 (Syt1(-/-), Syt2(-/-) and Syt7(-/-) mice). We only found a defect in RRP recovery in Syt1(-/-) mice which was apparent as a strongly reduced response to repetitive stimulations. In post-hearing Syt2(-/-) and Syt7(-/-) mutant mice, IHC synaptic exocytosis was unaffected. The transient expression of Syt1 and Syt2, which were no longer detected in IHCs after the onset of hearing, indicates that these two most common Ca(2+)-sensors in CNS synapses are not involved in mature IHCs. We suggest that otoferlin underlies highly efficient Ca(2+)-dependent membrane-membrane fusion, a process likely essential to increase the probability and synchrony of vesicle fusion events at the mature IHC ribbon synapse.
Figures
Comment in
-
Deciphering the roles of C(2)-domain-containing proteins (synaptotagmins and otoferlin) in the inner ear.J Neurosci. 2011 Mar 30;31(13):4765-7. doi: 10.1523/JNEUROSCI.6359-10.2011. J Neurosci. 2011. PMID: 21451013 Free PMC article. No abstract available.
References
-
- Achanzar WE, Ward S. A nematode gene required for sperm vesicle fusion. J Cell Sci. 1997;110:1073–1081. - PubMed
-
- Bansal D, Campbell KP. Dysferlin and the plasma membrane repair in muscular dystrophy. Trends Cell Biol. 2004;14:206–213. - PubMed
-
- Bansal D, Miyake K, Vogel SS, Groh S, Chen CC, Williamson R, McNeil PL, Campbell KP. Defective membrane repair in dysferlin-deficient muscular dystrophy. Nature. 2003;423:168–172. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous