

Quantitative analysis of synaptic vesicle Rabs uncovers distinct yet overlapping roles for Rab3a and Rab27b in Ca2+-triggered exocytosis
- PMID: 20926670
- PMCID: PMC6634719
- DOI: 10.1523/JNEUROSCI.0907-10.2010
Quantitative analysis of synaptic vesicle Rabs uncovers distinct yet overlapping roles for Rab3a and Rab27b in Ca2+-triggered exocytosis
Abstract
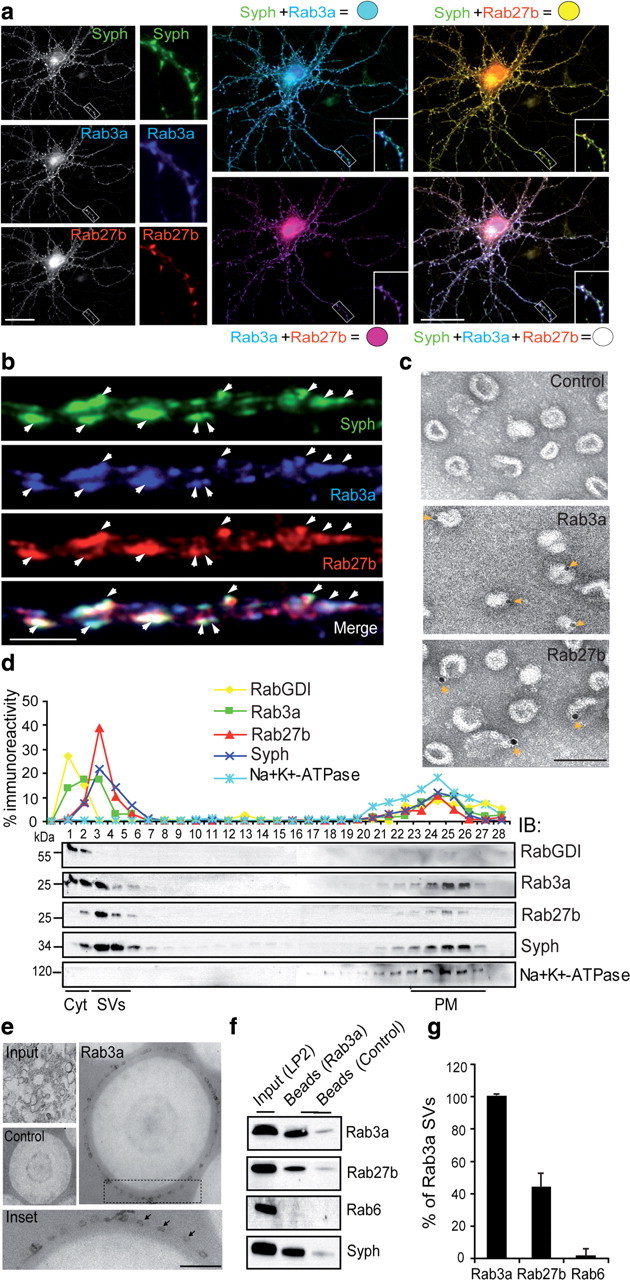
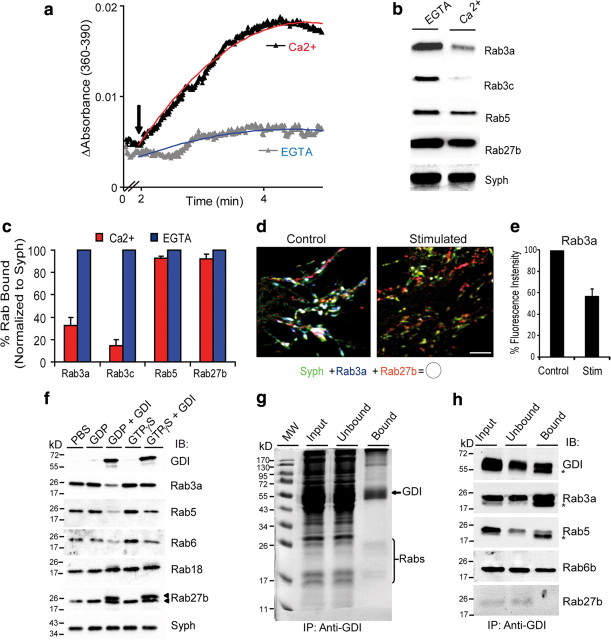
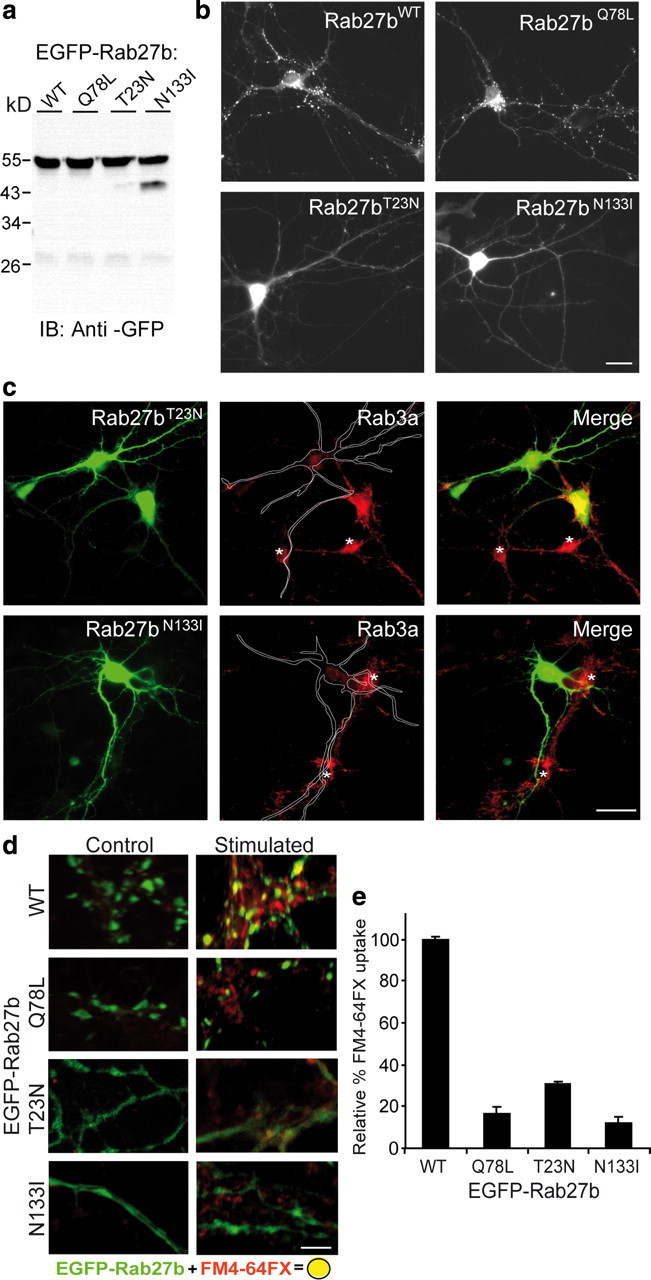
Rab GTPases are molecular switches that orchestrate protein complexes before membrane fusion reactions. In synapses, Rab3 and Rab5 proteins have been implicated in the exo-endocytic cycling of synaptic vesicles (SVs), but an involvement of additional Rabs cannot be excluded. Here, combining high-resolution mass spectrometry and chemical labeling (iTRAQ) together with quantitative immunoblotting and fluorescence microscopy, we have determined the exocytotic (Rab3a, Rab3b, Rab3c, and Rab27b) and endocytic (Rab4b, Rab5a/b, Rab10, Rab11b, and Rab14) Rab machinery of SVs. Analysis of two closely related proteins, Rab3a and Rab27b, revealed colocalization in synaptic nerve terminals, where they reside on distinct but overlapping SV pools. Moreover, whereas Rab3a readily dissociates from SVs during Ca(2+)-triggered exocytosis, and is susceptible to membrane extraction by Rab-GDI, Rab27b persists on SV membranes upon stimulation and is resistant to GDI-coupled Rab retrieval. Finally, we demonstrate that selective modulation of the GTP/GDP switch mechanism of Rab27b impairs SV recycling, suggesting that Rab27b, probably in concert with Rab3s, is involved in SV exocytosis.
Figures




References
-
- Burstein ES, Brondyk WH, Macara IG. Amino acid residues in the Ras-like GTPase Rab3A that specify sensitivity to factors that regulate the GTP/GDP cycling of Rab3A. J Biol Chem. 1992;267:22715–22718. - PubMed
-
- Castillo PE, Schoch S, Schmitz F, Südhof TC, Malenka RC. RIM1alpha is required for presynaptic long-term potentiation. Nature. 2002;415:327–330. - PubMed
-
- Chavas LM, Torii S, Kamikubo H, Kawasaki M, Ihara K, Kato R, Kataoka M, Izumi T, Wakatsuki S. Structure of the small GTPase Rab27b shows an unexpected swapped dimer. Acta Crystallogr D Biol Crystallogr. 2007;63:769–779. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous