

β1A integrin is a master regulator of invadosome organization and function
- PMID: 20926684
- PMCID: PMC2993740
- DOI: 10.1091/mbc.E10-07-0580
β1A integrin is a master regulator of invadosome organization and function
Abstract
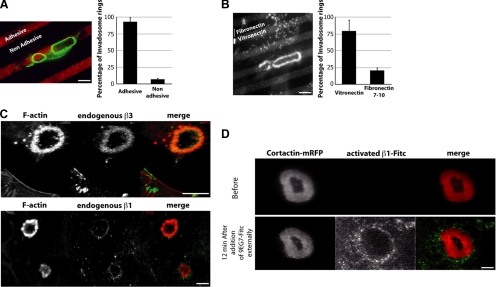
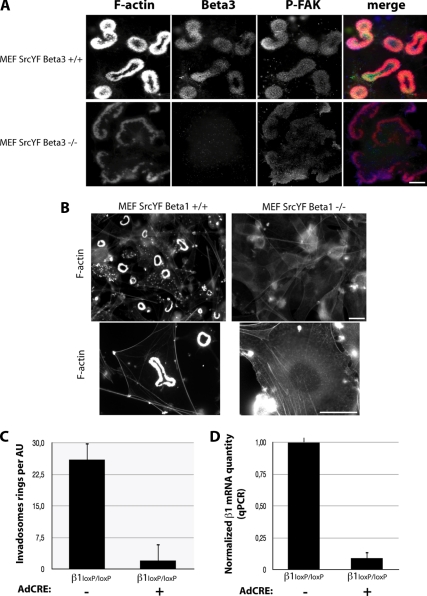
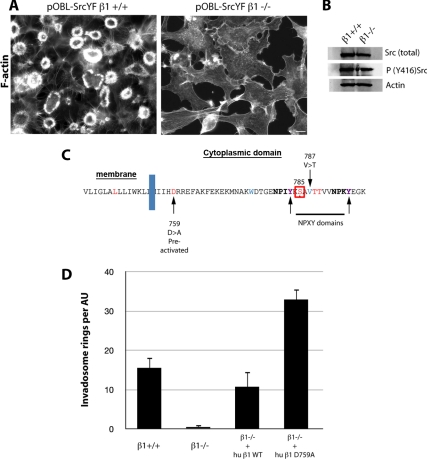
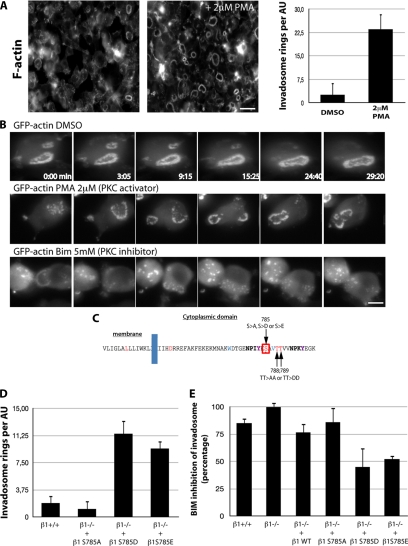
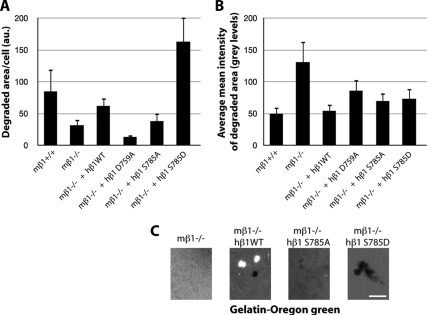
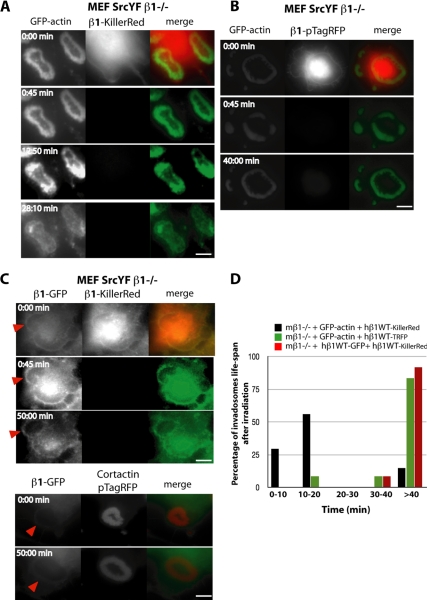
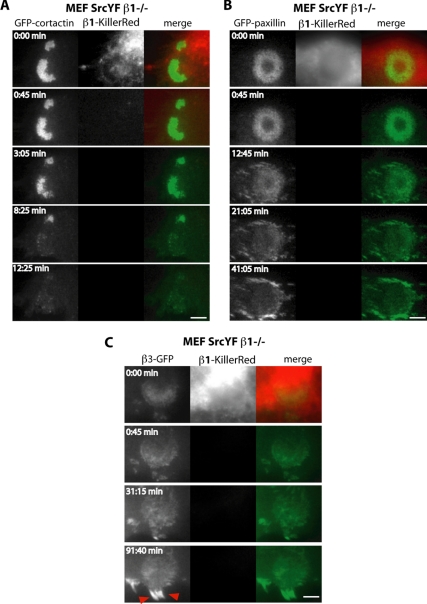
Invadosomes are adhesion structures involved in tissue invasion that are characterized by an intense actin polymerization-depolymerization associated with β1 and β3 integrins and coupled to extracellular matrix (ECM) degradation activity. We induced the formation of invadosomes by expressing the constitutive active form of Src, SrcYF, in different cell types. Use of ECM surfaces micropatterned at the subcellular scale clearly showed that in mesenchymal cells, integrin signaling controls invadosome activity. Using β1⁻/⁻ or β3⁻/⁻ cells, it seemed that β1A but not β3 integrins are essential for initiation of invadosome formation. Protein kinase C activity was shown to regulate autoassembly of invadosomes into a ring-like metastructure (rosette), probably by phosphorylation of Ser785 on the β1A tail. Moreover, our study clearly showed that β1A links actin dynamics and ECM degradation in invadosomes. Finally, a new strategy based on fusion of the photosensitizer KillerRed to the β1A cytoplasmic domain allowed specific and immediate loss of function of β1A, resulting in disorganization and disassembly of invadosomes and formation of focal adhesions.
Figures
References
-
- Artym V. V., Zhang Y., Seillier-Moiseiwitsch F., Yamada K. M., Mueller S. C. Dynamic interactions of cortactin and membrane type 1 matrix metalloproteinase at invadopodia: defining the stages of invadopodia formation and function. Cancer Res. 2006;66:3034–3043. - PubMed
-
- Ayala I., Baldassarre M., Giacchetti G., Caldieri G., Tete S., Luini A., Buccione R. Multiple regulatory inputs converge on cortactin to control invadopodia biogenesis and extracellular matrix degradation. J. Cell Sci. 2008;121:369–378. - PubMed
-
- Bellows C. G., Sodek J., Yao K. L., Aubin J. E. Phenotypic differences in subclones and long-term cultures of clonally derived rat bone cell lines. J. Cell. Biochem. 1986;31:153–169. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
