

ATF3, an adaptive-response gene, enhances TGF{beta} signaling and cancer-initiating cell features in breast cancer cells
- PMID: 20930144
- PMCID: PMC2951469
- DOI: 10.1242/jcs.064915
ATF3, an adaptive-response gene, enhances TGF{beta} signaling and cancer-initiating cell features in breast cancer cells
Abstract
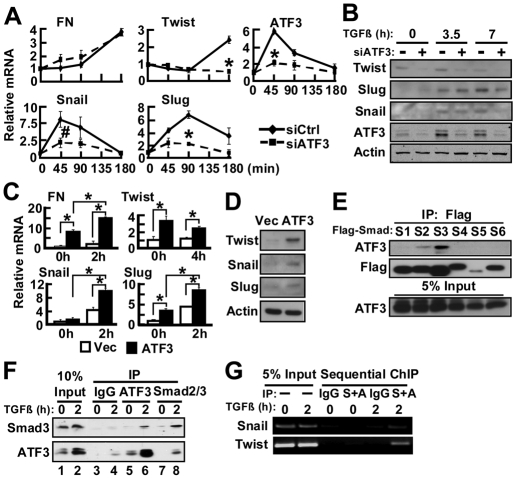
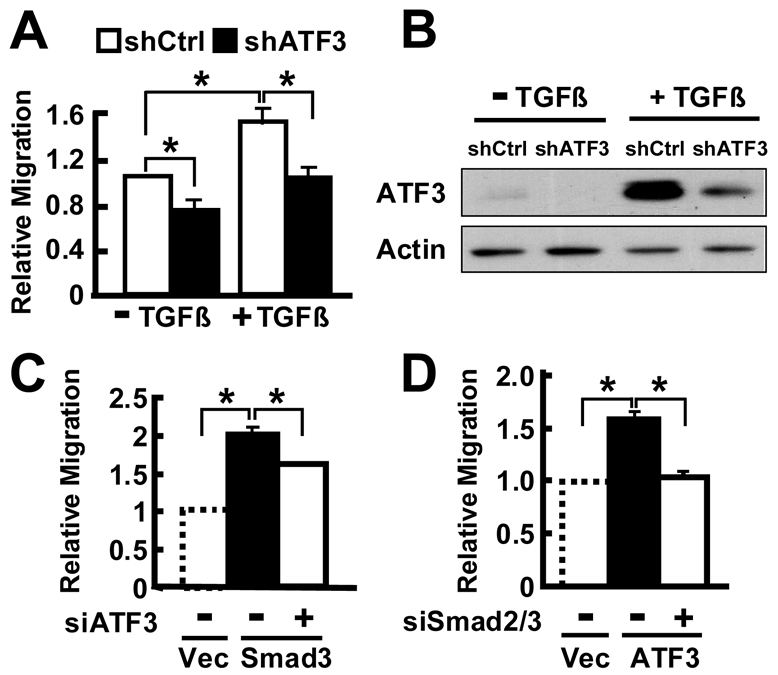
The activating transcription factor 3 (ATF3) gene is induced by a variety of signals, including many of those encountered by cancer cells. We present evidence that ATF3 is induced by TGFβ in the MCF10CA1a breast cancer cells and plays an integral role for TGFβ to upregulate its target genes snail, slug and twist, and to enhance cell motility. Furthermore, ATF3 upregulates the expression of the TGFb gene itself, forming a positive-feedback loop for TGFβ signaling. Functionally, ectopic expression of ATF3 leads to morphological changes and alterations of markers consistent with epithelial-to-mesenchymal transition (EMT). It also leads to features associated with breast-cancer-initiating cells: increased CD24(low)-CD44(high) population of cells, mammosphere formation and tumorigenesis. Conversely, knockdown of ATF3 reduces EMT, CD24(low)-CD44(high) cells and mammosphere formation. Importantly, knocking down twist, a downstream target, reduces the ability of ATF3 to enhance mammosphere formation, indicating the functional significance of twist in ATF3 action. To our knowledge, this is the first report demonstrating the ability of ATF3 to enhance breast cancer-initiating cell features and to feedback on TGFβ. Because ATF3 is an adaptive-response gene and is induced by various stromal signals, these findings have significant implications for how the tumor microenvironment might affect cancer development.
Figures




Similar articles
-
SLUG/SNAI2 and tumor necrosis factor generate breast cells with CD44+/CD24- phenotype.BMC Cancer. 2010 Aug 6;10:411. doi: 10.1186/1471-2407-10-411. BMC Cancer. 2010. PMID: 20691079 Free PMC article.
-
PARP3 controls TGFβ and ROS driven epithelial-to-mesenchymal transition and stemness by stimulating a TG2-Snail-E-cadherin axis.Oncotarget. 2016 Sep 27;7(39):64109-64123. doi: 10.18632/oncotarget.11627. Oncotarget. 2016. PMID: 27579892 Free PMC article.
-
Twist modulates breast cancer stem cells by transcriptional regulation of CD24 expression.Neoplasia. 2009 Dec;11(12):1318-28. doi: 10.1593/neo.91084. Neoplasia. 2009. PMID: 20019840 Free PMC article.
-
Reprogramming during epithelial to mesenchymal transition under the control of TGFβ.Cell Adh Migr. 2015;9(3):233-46. doi: 10.4161/19336918.2014.983794. Epub 2014 Nov 17. Cell Adh Migr. 2015. PMID: 25482613 Free PMC article. Review.
-
Epithelial mesenchymal transition traits in human breast cancer cell lines parallel the CD44(hi/)CD24 (lo/-) stem cell phenotype in human breast cancer.J Mammary Gland Biol Neoplasia. 2010 Jun;15(2):235-52. doi: 10.1007/s10911-010-9175-z. Epub 2010 Jun 4. J Mammary Gland Biol Neoplasia. 2010. PMID: 20521089 Review.
Cited by
-
Overexpression of activating transcription factor 3 exerts suppressive effects in HepG2 cells.Mol Med Rep. 2019 Feb;19(2):869-876. doi: 10.3892/mmr.2018.9707. Epub 2018 Nov 29. Mol Med Rep. 2019. PMID: 30535500 Free PMC article.
-
The antioxidant transcription factor Nrf2 modulates the stress response and phenotype of malignant as well as premalignant pancreatic ductal epithelial cells by inducing expression of the ATF3 splicing variant ΔZip2.Oncogene. 2019 Feb;38(9):1461-1476. doi: 10.1038/s41388-018-0518-3. Epub 2018 Oct 9. Oncogene. 2019. PMID: 30302023
-
ATF3, a hub of the cellular adaptive-response network, in the pathogenesis of diseases: is modulation of inflammation a unifying component?Gene Expr. 2010;15(1):1-11. doi: 10.3727/105221610x12819686555015. Gene Expr. 2010. PMID: 21061913 Free PMC article. Review.
-
HER2 Upregulates ATF4 to Promote Cell Migration via Activation of ZEB1 and Downregulation of E-Cadherin.Int J Mol Sci. 2019 May 6;20(9):2223. doi: 10.3390/ijms20092223. Int J Mol Sci. 2019. PMID: 31064130 Free PMC article.
-
Runx2, a target gene for activating transcription factor-3 in human breast cancer cells.Tumour Biol. 2015 Mar;36(3):1923-31. doi: 10.1007/s13277-014-2796-x. Epub 2014 Nov 8. Tumour Biol. 2015. PMID: 25380580
References
-
- Abe M., Harpel J. G., Metz C. N., Nunes I., Loskutoff D. J., Rifkin D. B. (1994). An assay for transforming growth factor-beta using cells transfected with a plasminogen activator inhibitor-1 promoter-luciferase construct. Anal. Biochem. 216, 276-284 - PubMed
-
- Ailles L. E., Weissman I. L. (2007). Cancer stem cells in solid tumors. Curr. Opin. Biotechnol. 18, 460-466 - PubMed
-
- Ameri K., Hammond E. M., Culmsee C., Raida M., Katschinski D. M., Wenger R. H., Wagner E., Davis R. J., Hai T., Denko N., et al. (2007). Induction of activating transcription factor 3 by anoxia is independent of p53 and the hypoxic HIF signalling pathway. Oncogene 26, 284-289 - PubMed
-
- Bandyopadhyay S., Wang Y., Zhan R., Pai S. K., Watabe M., Iiizumi M., Furuta E., Mohinta S., Liu W., Hirota S., et al. (2006). The tumor metastasis suppressor gene Drg-1 down regulates the expression of ATF3 in prostate cancer. Can. Res. 66, 11983-11990 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
