

Ribozymes and riboswitches: modulation of RNA function by small molecules
- PMID: 20931966
- PMCID: PMC2965775
- DOI: 10.1021/bi1012645
Ribozymes and riboswitches: modulation of RNA function by small molecules
Abstract
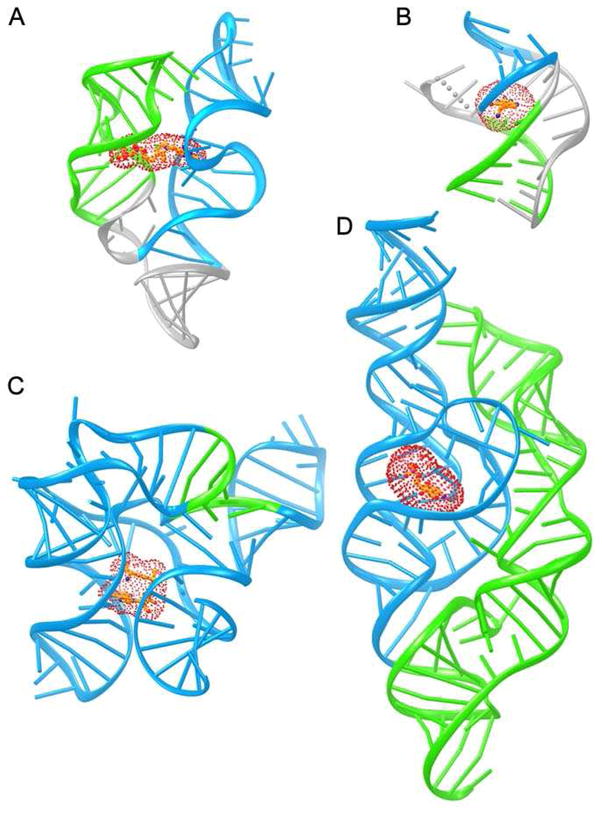
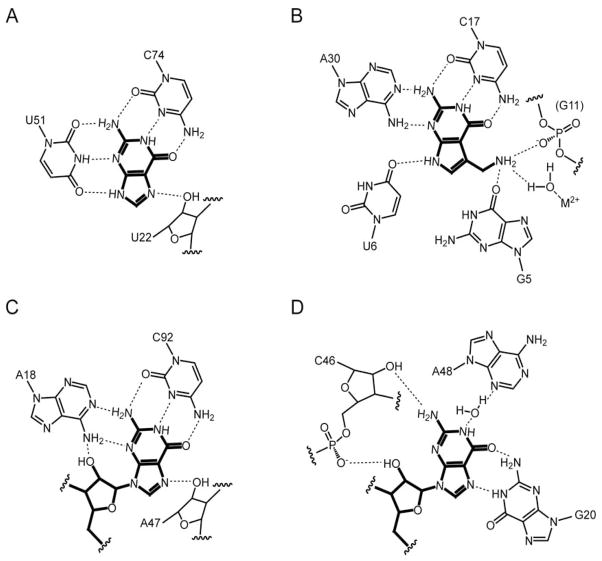
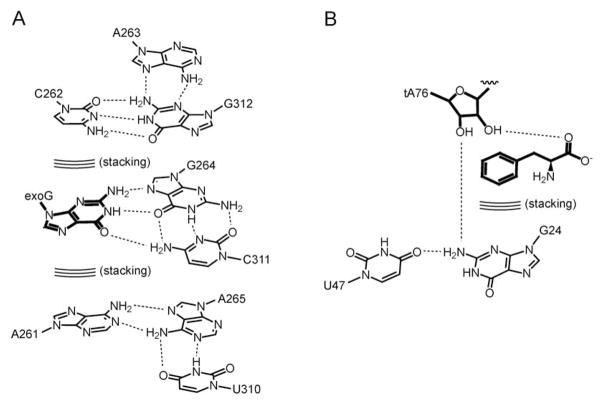
Diverse small molecules interact with catalytic RNAs (ribozymes) as substrates and cofactors, and their intracellular concentrations are sensed by gene-regulatory mRNA domains (riboswitches) that modulate transcription, splicing, translation, or RNA stability. Although recognition mechanisms vary from RNA to RNA, structural analyses reveal recurring strategies that arise from the intrinsic properties of RNA such as base pairing and stacking with conjugated heterocycles, and cation-dependent recognition of anionic functional groups. These studies also suggest that, to a first approximation, the magnitude of ligand-induced reorganization of an RNA is inversely proportional to the complexity of the riboswitch or ribozyme. How these small molecule binding-induced changes in RNA lead to alteration in gene expression is less well understood. While different riboswitches have been proposed to be under either kinetic or thermodynamic control, the biochemical and structural mechanisms that give rise to regulatory consequences downstream of small molecule recognition by RNAs mostly remain to be elucidated.
Figures


Similar articles
-
A structural analysis of in vitro catalytic activities of hammerhead ribozymes.BMC Bioinformatics. 2007 Nov 30;8:469. doi: 10.1186/1471-2105-8-469. BMC Bioinformatics. 2007. PMID: 18053134 Free PMC article.
-
The dynamic nature of RNA as key to understanding riboswitch mechanisms.Acc Chem Res. 2011 Dec 20;44(12):1339-48. doi: 10.1021/ar200035g. Epub 2011 Jun 16. Acc Chem Res. 2011. PMID: 21678902
-
Fundamental studies of functional nucleic acids: aptamers, riboswitches, ribozymes and DNAzymes.Chem Soc Rev. 2020 Oct 21;49(20):7331-7353. doi: 10.1039/d0cs00617c. Epub 2020 Sep 18. Chem Soc Rev. 2020. PMID: 32944725 Review.
-
Riboswitches: discovery of drugs that target bacterial gene-regulatory RNAs.Acc Chem Res. 2011 Dec 20;44(12):1329-38. doi: 10.1021/ar200039b. Epub 2011 May 26. Acc Chem Res. 2011. PMID: 21615107 Free PMC article. Review.
-
RNA-based networks: using RNA aptamers and ribozymes as synthetic genetic devices.Methods Mol Biol. 2012;813:157-68. doi: 10.1007/978-1-61779-412-4_9. Methods Mol Biol. 2012. PMID: 22083741
Cited by
-
Basis for ligand discrimination between ON and OFF state riboswitch conformations: the case of the SAM-I riboswitch.RNA. 2012 Jun;18(6):1230-43. doi: 10.1261/rna.032177.111. Epub 2012 Apr 27. RNA. 2012. PMID: 22543867 Free PMC article.
-
RNA motifs and combinatorial prediction of interactions, stability and localization of noncoding RNAs.Nat Struct Mol Biol. 2018 Dec;25(12):1070-1076. doi: 10.1038/s41594-018-0155-0. Epub 2018 Nov 12. Nat Struct Mol Biol. 2018. PMID: 30420773 Review.
-
Rapid RNA-ligand interaction analysis through high-information content conformational and stability landscapes.Nat Commun. 2015 Dec 7;6:8898. doi: 10.1038/ncomms9898. Nat Commun. 2015. PMID: 26638992 Free PMC article.
-
A Two-Way Street: Regulatory Interplay between RNA Polymerase and Nascent RNA Structure.Trends Biochem Sci. 2016 Apr;41(4):293-310. doi: 10.1016/j.tibs.2015.12.009. Epub 2016 Jan 25. Trends Biochem Sci. 2016. PMID: 26822487 Free PMC article. Review.
-
Parallel Discovery Strategies Provide a Basis for Riboswitch Ligand Design.Cell Chem Biol. 2020 Oct 15;27(10):1241-1249.e4. doi: 10.1016/j.chembiol.2020.07.021. Epub 2020 Aug 13. Cell Chem Biol. 2020. PMID: 32795418 Free PMC article.
References
-
- Loh E, Dussurget O, Gripenland J, Vaitkevicius K, Tiensuu T, Mandin P, Repoila F, Buchrieser C, Cossart P, Johansson J. A trans-acting riboswitch controls expression of the virulence regulator PrfA in Listeria monocytogenes. Cell. 2010;139:770–779. - PubMed
-
- Rajkowitsch L, Chen D, Stampfl S, Semrad K, Waldsich C, Mayer O, Jantsch MF, Konrat R, Bläsi U, Schroeder R. RNA chaperones, RNA annealers and RNA helicases. RNA Biology. 2007;4:118–130. - PubMed
-
- Winkler W, Nahvi A, Breaker RR. Thiamine derivatives bind messenger RNAs directly to regulate bacterial gene expression. Nature. 2002;419:952–956. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
