

Mammalian target of rapamycin controls dendritic cell development downstream of Flt3 ligand signaling
- PMID: 20933441
- PMCID: PMC2966531
- DOI: 10.1016/j.immuni.2010.09.012
Mammalian target of rapamycin controls dendritic cell development downstream of Flt3 ligand signaling
Abstract
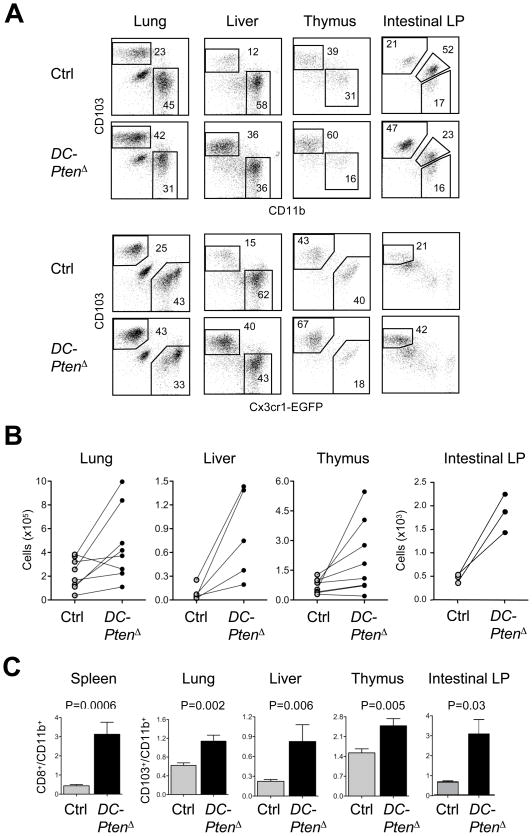
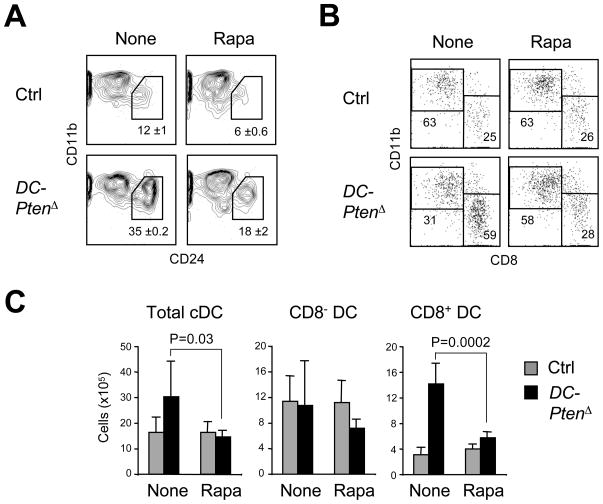
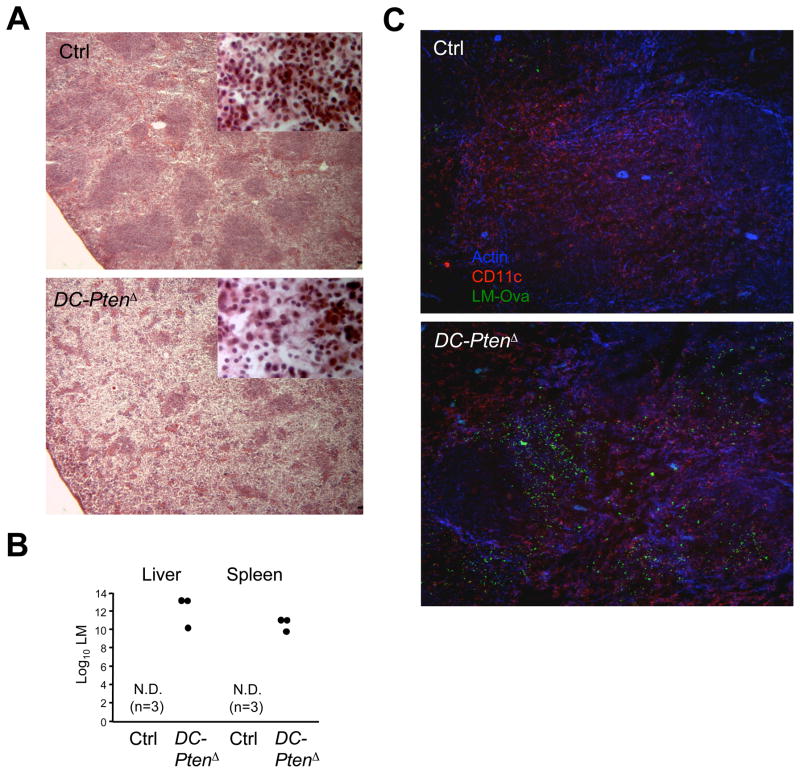
Dendritic cells (DCs) comprise distinct functional subsets including CD8⁻ and CD8(+) classical DCs (cDCs) and interferon-secreting plasmacytoid DCs (pDCs). The cytokine Flt3 ligand (Flt3L) controls the development of DCs and is particularly important for the pDC and CD8(+) cDC and their CD103(+) tissue counterparts. We report that mammalian target of rapamycin (mTOR) inhibitor rapamycin impaired Flt3L-driven DC development in vitro, with the pDCs and CD8(+)-like cDCs most profoundly affected. Conversely, deletion of the phosphoinositide 3-kinase (PI3K)-mTOR negative regulator Pten facilitated Flt3L-driven DC development in culture. DC-specific Pten targeting in vivo caused the expansion of CD8(+) and CD103(+) cDC numbers, which was reversible by rapamycin. The increased CD8(+) cDC numbers caused by Pten deletion correlated with increased susceptibility to the intracellular pathogen Listeria. Thus, PI3K-mTOR signaling downstream of Flt3L controls DC development, and its restriction by Pten ensures optimal DC pool size and subset composition.
Copyright © 2010 Elsevier Inc. All rights reserved.
Conflict of interest statement
Technologies associated with phospho-flow are licensed in part to BD Biosciences, and G.P.N. is a consultant for BD Biosciences, a supplier of the reagents used in this report. Other authors declare no competing interests.
Figures
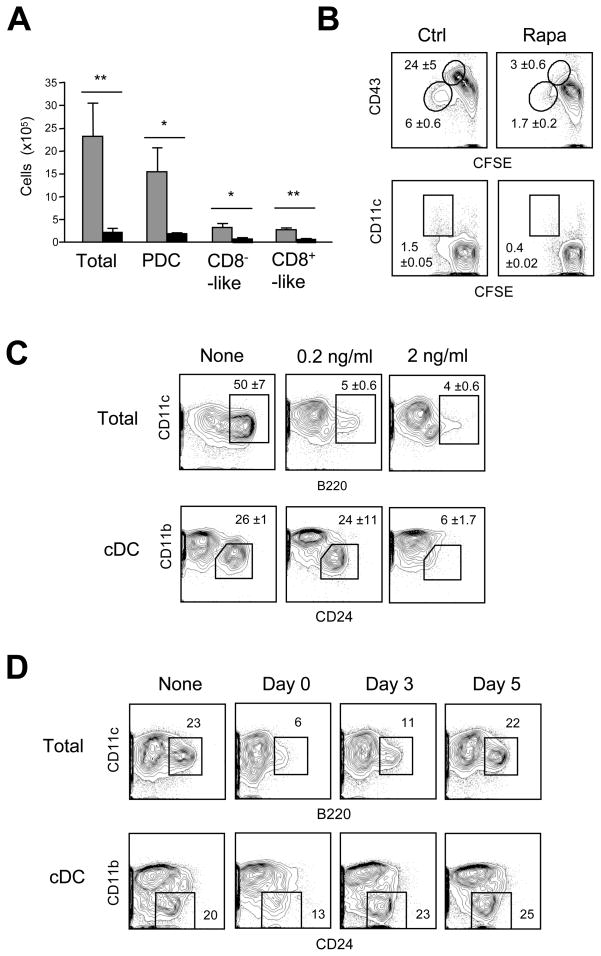
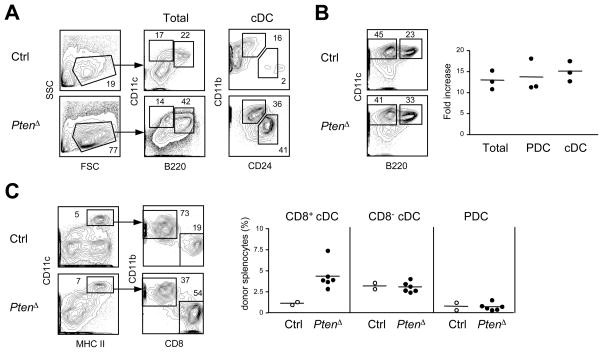
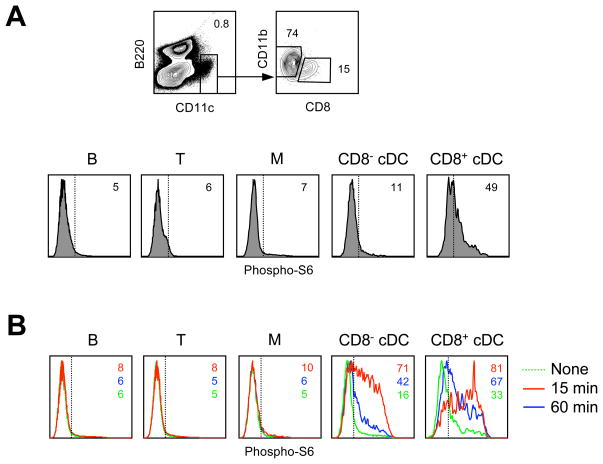
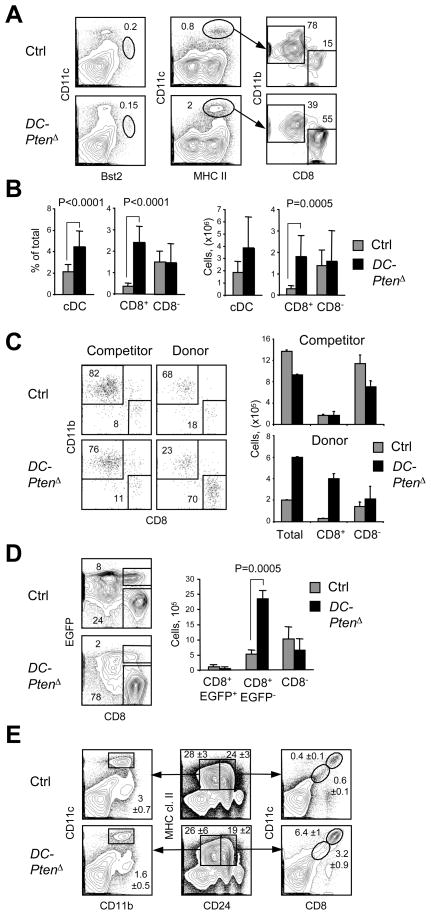
Comment in
-
A Flt3L encounter: mTOR signaling in dendritic cells.Immunity. 2010 Oct 29;33(4):580-2. doi: 10.1016/j.immuni.2010.10.001. Immunity. 2010. PMID: 21029968
References
-
- Alaniz RC, Sandall S, Thomas EK, Wilson CB. Increased dendritic cell numbers impair protective immunity to intracellular bacteria despite augmenting antigen-specific CD8+ T lymphocyte responses. J Immunol. 2004;172:3725–3735. - PubMed
-
- Anzelon AN, Wu H, Rickert RC. Pten inactivation alters peripheral B lymphocyte fate and reconstitutes CD19 function. Nat Immunol. 2003;4:287–294. - PubMed
-
- Aoshi T, Zinselmeyer BH, Konjufca V, Lynch JN, Zhang X, Koide Y, Miller MJ. Bacterial entry to the splenic white pulp initiates antigen presentation to CD8+ T cells. Immunity. 2008;29:476–486. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
