

Emergence of Pseudomonas aeruginosa strains producing high levels of persister cells in patients with cystic fibrosis
- PMID: 20935098
- PMCID: PMC2981199
- DOI: 10.1128/JB.01651-09
Emergence of Pseudomonas aeruginosa strains producing high levels of persister cells in patients with cystic fibrosis
Abstract
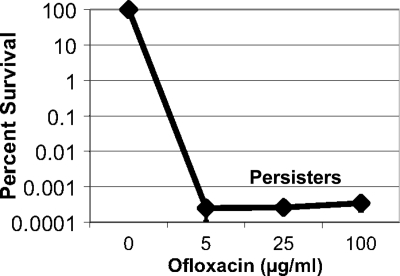
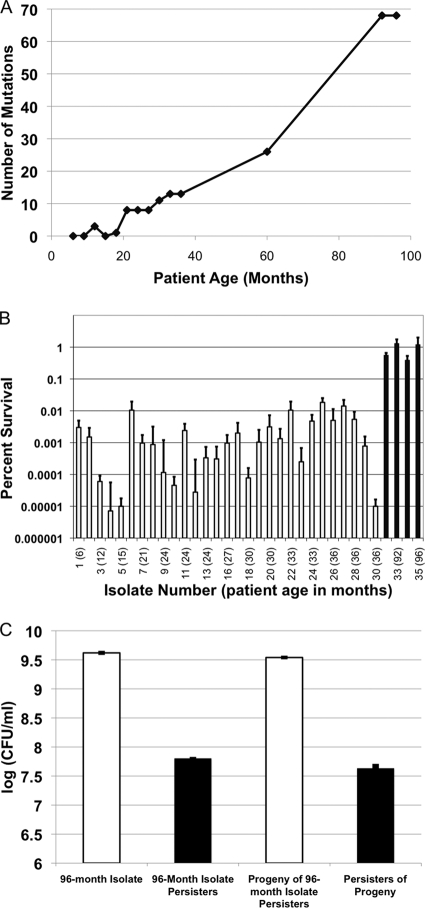
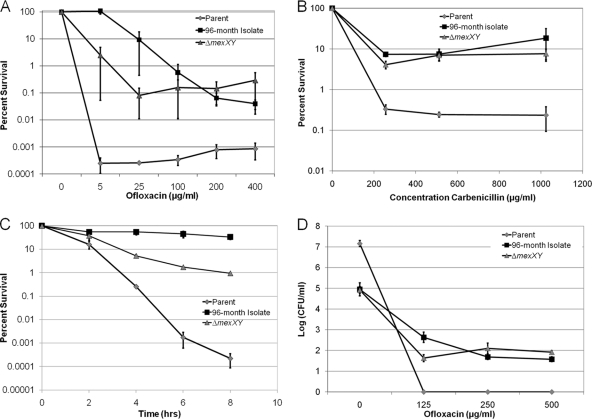
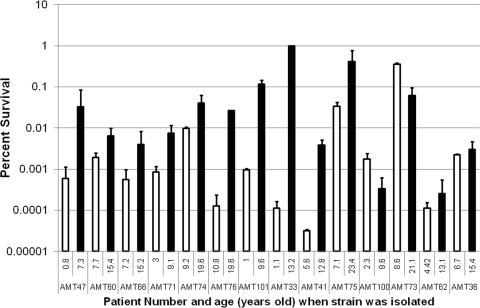
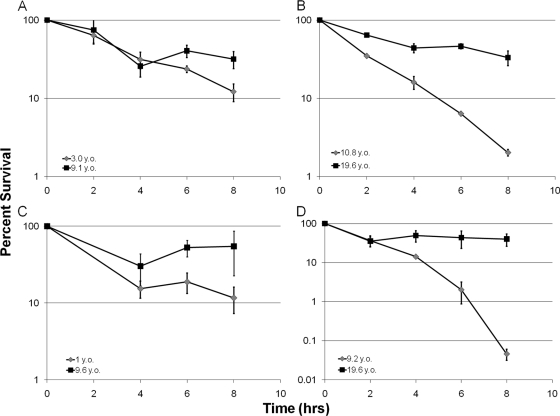
The majority of cystic fibrosis (CF) patients succumb to a chronic infection of the airway with Pseudomonas aeruginosa. Paradoxically, pathogenic strains are often susceptible to antibiotics, but the infection cannot be eradicated with antimicrobial therapy. We find that in a majority of patients with airway infections, late isolates of P. aeruginosa produce increased levels of drug-tolerant persister cells. The genomes of a clonal pair of early/late isolates from a single patient have been previously sequenced, and the late isolate (obtained at age 96 months) showed a 100-fold increase in persister levels. The 96-month isolate carries a large number of mutations, including a mutation in mutS that confers a hypermutator phenotype. There is also a mutation in the mexZ repressor controlling the expression of the MexXY-OprM multidrug pump, which results in a moderate increase in the ofloxacin, carbenicillin, and tobramycin MICs. Knocking out the mexXY locus restored the resistance to that of the parent strain but did not affect the high levels of persisters formed by the 96-month isolate. This suggests that the late isolate is a high-persister (hip) mutant. Increased persister formation was observed in exponential phase, stationary phase, and biofilm populations of the 96-month isolate. Analysis of late isolates from 14 additional patients indicated that 10 of them are hip mutants. Most of these hip mutants did not have higher drug resistance. Increased persister formation appears to be their sole mechanism for surviving chemotherapy. Taken together, these findings suggest a link between persisters and recalcitrance of CF infection and identify an overlooked culprit-high-persister mutants producing elevated levels of drug-tolerant cells. Persisters may play a similarly critical role in the recalcitrance of other chronic infections.
Figures
References
-
- Alhede, M., T. Bjarnsholt, P. O. Jensen, R. K. Phipps, C. Moser, L. Christophersen, L. D. Christensen, M. van Gennip, M. Parsek, N. Hoiby, T. B. Rasmussen, and M. Givskov. 2009. Pseudomonas aeruginosa recognizes and responds aggressively to the presence of polymorphonuclear leukocytes. Microbiology 155:3500-3508. - PubMed
-
- Bjarnsholt, T., P. Ø. Jensen, M. J. Fiandaca, J. Pedersen, C. R. Hansen, C. B. Andersen, T. Pressler, M. Givskov, and N. Høiby. 2009. Pseudomonas aeruginosa biofilms in the respiratory tract of cystic fibrosis patients. Pediatr. Pulmonol. 44:547-558. - PubMed
-
- Bjarnsholt, T., K. Kirketerp-Moller, P. O. Jensen, K. G. Madsen, R. Phipps, K. Krogfelt, N. Hoiby, and M. Givskov. 2008. Why chronic wounds will not heal: a novel hypothesis. Wound Repair Regen. 16:2-10. - PubMed
-
- Burns, J. L., J. M. Van Dalfsen, R. M. Shawar, K. L. Otto, R. L. Garber, J. M. Quan, A. B. Montgomery, G. M. Albers, B. W. Ramsey, and A. L. Smith. 1999. Effect of chronic intermittent administration of inhaled tobramycin on respiratory microbial flora in patients with cystic fibrosis. J. Infect. Dis. 179:1190-1196. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
