

doi: 10.4061/2010/389129.
Nonhomologous DNA end joining in cell-free extracts
Affiliations
- PMID: 20936167
- PMCID: PMC2945661
- DOI: 10.4061/2010/389129
Item in Clipboard
Nonhomologous DNA end joining in cell-free extracts
J Nucleic Acids.
.
Abstract
Among various DNA damages, double-strand breaks (DSBs) are considered as most deleterious, as they may lead to chromosomal rearrangements and cancer when unrepaired. Nonhomologous DNA end joining (NHEJ) is one of the major DSB repair pathways in higher organisms. A large number of studies on NHEJ are based on in vitro systems using cell-free extracts. In this paper, we summarize the studies on NHEJ performed by various groups in different cell-free repair systems.
Figures
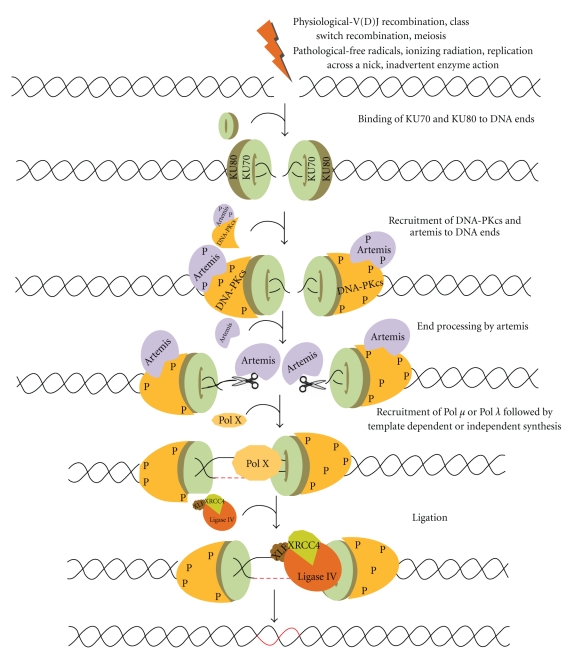
Double-strand breaks (DSBs) are generated exogenously by ionizing radiation, or endogenously by free radicals or during V(D)J recombination in pre-B (bone marrow) and pre-T cells (thymus) by RAG complex or also during class switch recombination in activated B cells (in the peripheral lymphoid tissues such as spleen, lymph nodes, and Peyer's patches). NHEJ involves the binding of Ku70 and Ku80 heterodimeric complex to the DNA ends, and DNA-PKcs in association with ARTEMIS. ARTEMIS is a 5′-3′ exonuclease in an unphosphorylated form while it is an endonuclease in a phosphorylated form. Artemis protein acts as an exonuclease and helps in resection of the ends. Polymerase X family members are then recruited for DNA synthesis, which includes both template dependent and independent DNA synthesis. The resulting DNA ends are ligated by a specific DNA LIGASE IV with stimulatory factors (XRCC4-LIGASE IV-XLF complex) that restores the integrity of DNA.
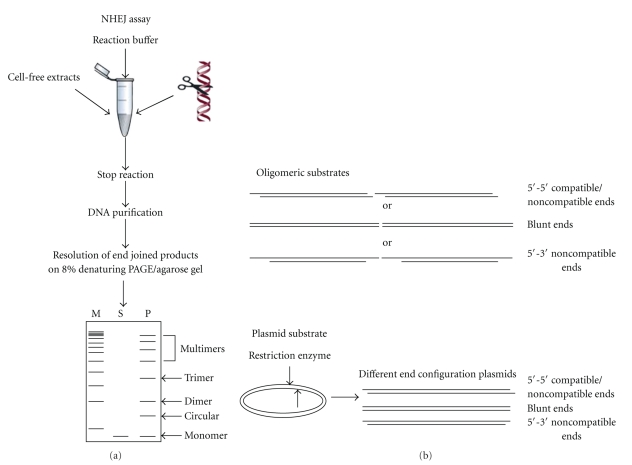
Schematic representation of NHEJ reaction using cell-free extracts. (a) Cell-free extract is incubated with one of the 5′-end-labelled DNA substrate (oligomer) or restriction enzyme-digested plasmid DNA as shown in the figure along with NHEJ buffer. The reaction is stopped by adding proteinase K and EDTA, DNA is purified by phenol:chloroform extraction, precipitated with glycogen and ethanol, resuspended in TE, and run on 8% denaturing PAGE or agarose gel. After end joining, dimers, trimers, and multimers can be seen. “M” is marker, “S” is substrate, and “P” indicates products. (b) Oligomeric or plasmid DNA substrates
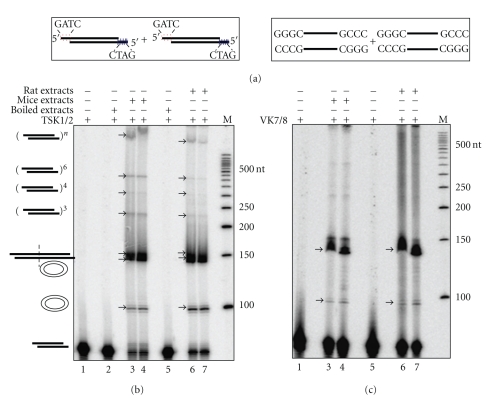
Comparison of efficiencies of NHEJ in rat and mice testis. Cell-free extract was prepared from age-matched rat and mice testes and protein profile was normalized between both animals. 5 μg of protein was incubated with 4 nM of 5′-end labelled with [γ
−32P] DNA containing both compatible and blunt termini against each lane (a) which are represented schematically along with buffer containing 30 mM HEPES-KOH, pH 7.9, 7.5 mM MgCl2, 1 mM DTT, 2 mM ATP, 50 μM dNTPs, and 0.1 μg BSA. End joining reaction using (b) compatible end and (c) blunt end DNA. Lane 1 shows negative control that contains substrate alone, Lane 2 shows heat-inactivated control which is mice testis cell-free extract-boiled for 10 min and used for the reaction, and Lanes 3 and 4 are the end joining reactions with cell-free extract from mice testis. Lane 5 is the heat-inactivated control which is boiled rat testicular extract as described previously. Lanes 6 and 7 are the end joining reactions with rat testicular cell-free extracts. “M” indicates 5′-end-labelled 50 bp ladder. The efficiency of joining is similar in both mice and rat testicular extracts. Different types of end-joined products formed are indicated.

Schematic representation of possible mechanisms involved in repair for DNA termini. (a) In case of compatible ends, ligation usually does not involve much modification. Here, the two termini are aligned and ligated by DNA LIGASE IV. (b) Similarly, alignment of DNA ends in case of blunt ends take place and are ligated by DNA LIGASE IV as it does not need end modifications. On the other hand, modifications take place for non-compatible ends. There are different possibilities in case of 5′-5′ substrate which includes (c) deletion of nucleotides from both ends and insertion of nucleotides followed by ligation or (d) deletion of 5′-overhang from one end, end-filling of other 5′-end followed by blunt end ligation. (e) Another possibility includes end filling of both the overhangs followed by ligation. (f) In case of 5′-3′ overhangs, single strand ligation of overhangs is followed by template dependent synthesis and ligation. (g) Microhomology dependent joining can take place in any of the overhangs which is characterized by pairing of microhomology region followed by removal of flap by an endonuclease and then ligation.
References
-
- Lieber MR, Yu K, Raghavan SC. Roles of nonhomologous DNA end joining, V(D)J recombination, and class switch recombination in chromosomal translocations. DNA Repair. 2006;5(9-10):1234–1245. - PubMed
-
- Ferguson DO, Alt FW. DNA double strand break repair and chromosomal translocation: lessons from animal models. Oncogene. 2001;20(40):5572–5579. - PubMed
-
- Hefferin ML, Tomkinson AE. Mechanism of DNA double-strand break repair by non-homologous end joining. DNA Repair. 2005;4(6):639–648. - PubMed
-
- Weterings E, Chen DJ. The endless tale of non-homologous end-joining. Cell Research. 2008;18(1):114–124. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
